Эволюция. Почему глаза у нас спереди?
- Джейсон Г. Голдман
- BBC Future
Подпишитесь на нашу рассылку ”Контекст”: она поможет вам разобраться в событиях.
Автор фото, Thinkstock
Почему глаза у нас расположены не по бокам головы, а смотрят вперед? Отчасти это связано с необходимостью воспринимать трехмерные изображения, но корреспондент BBC Future обнаружил и другие причины.
Вы когда-нибудь обращали внимание, что большинство животных в зоопарке можно отнести к одной из двух групп? У одних глаза находятся по бокам головы (это куры, коровы, лошади, зебры), а у других они посажены ближе и расположены спереди (в эту группу входят обезьяны, тигры, совы и волки). Сами посетители зоопарка — люди — очевидно, относятся ко второй группе. С чем же связано это различие?
Расположение глаз — это всегда некий компромисс.
Автор фото, Thinkstock
Подпись к фото,У некоторых черепах глаза находятся по бокам, но мозг обрабатывает зрительную информацию так, как если бы глаза у них смотрели вперед
Вероятно, расположение глаз формировалось у разных животных по-разному. К примеру, у некоторых черепах глаза находятся по бокам, но мозг обрабатывает зрительную информацию так, как если бы глаза у них смотрели вперед, — возможно, это связано с тем, что когда черепахи втягивают голову под панцирь, их глаза воспринимают свет только спереди, как будто они расположены в передней части головы. Но почему у нашей ветви эволюционного древа — у приматов — глаза оказались спереди? Тому есть множество объяснений.
В 1922 году британский офтальмолог Эдвард Тричер Коллинз писал о том, что ранним приматам требовалось такое зрение, которое “позволяло бы им раскачиваться и точно перепрыгивать с ветки на ветку… хватать пищу руками и подносить ее ко рту”. Поэтому, решил ученый, в процессе эволюции у них развилась способность оценивать расстояние.
В последующие десятилетия гипотеза Коллинза неоднократно пересматривалась и уточнялась, но суть ее в течение долгого времени оставалась неизменной: в процессе эволюции глаза у наших предков переместились вперед, чтобы точно оценивать дистанцию при перепрыгивании с дерева на дерево. Цена ошибки при определении расстояния между деревьями действительно была немалой. “Расплатой за просчет было падение с высоты в несколько метров на землю, кишащую плотоядными зверями”, — написал в 1991 году специалист по визуальной психотерапии Кристофер Тайлер.
Автор фото, Thinkstock
Подпись к фото,Попугаи обладают панорамным зрением
Слабое место гипотезы Коллинза состоит в том, что у многих животных, которые селятся на деревьях, — например, у белок, — глаза расположены по бокам.
Американский нейробиолог Джон Оллман подхватил гипотезу Картмилла и доработал ее на основе сведений о ночных хищниках — ведь не у всех хищных животных глаза расположены спереди. У кошек, приматов и сов они действительно находятся в передней части головы, а у мангустов, тупай и мухоловок — по бокам.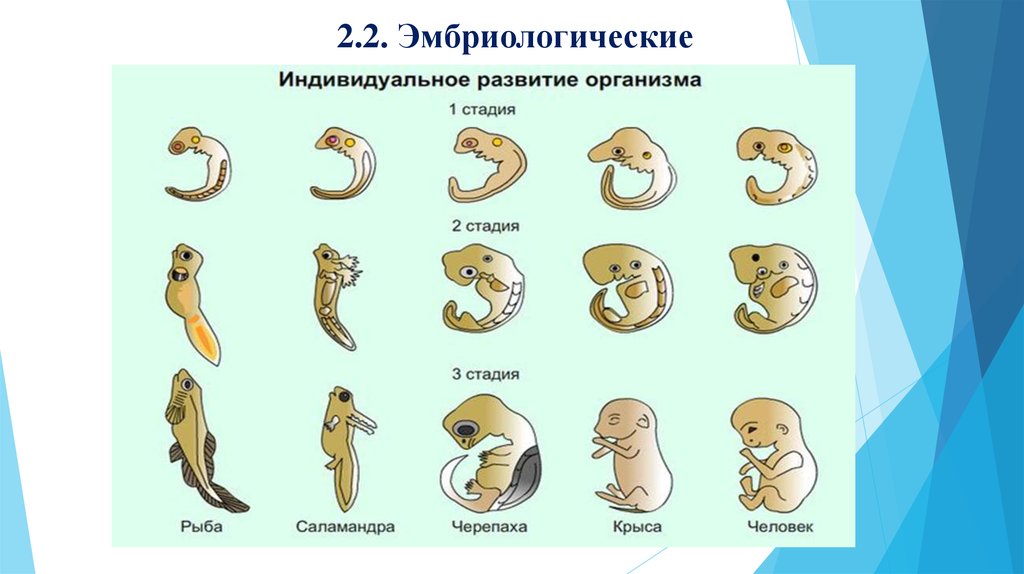
Автор фото, Thinkstock
Подпись к фото,У хищников, таких как этот леопард, глаза расположены спереди, чтобы лучше видеть добычу
У американского нейробиолога-теоретика Марка Чангизи возникло еще одно объяснение. В 2008 году он опубликовал в “Журнале теоретической биологии” (США) статью о “рентгеновском зрении”, предположив, что расположенные впереди глаза позволяли нашим предкам, жившим в лесу, видеть сквозь плотную листву и тесное переплетение веток. Громкое название “рентгеновское зрение” происходит от любопытного явления, описанного Чангизи: “Если держать палец перед глазами в вертикальном положении, фиксируя взгляд на каком-нибудь предмете, расположенном позади пальца, в мозг поступят два изображения пальца, и оба они будут прозрачными”.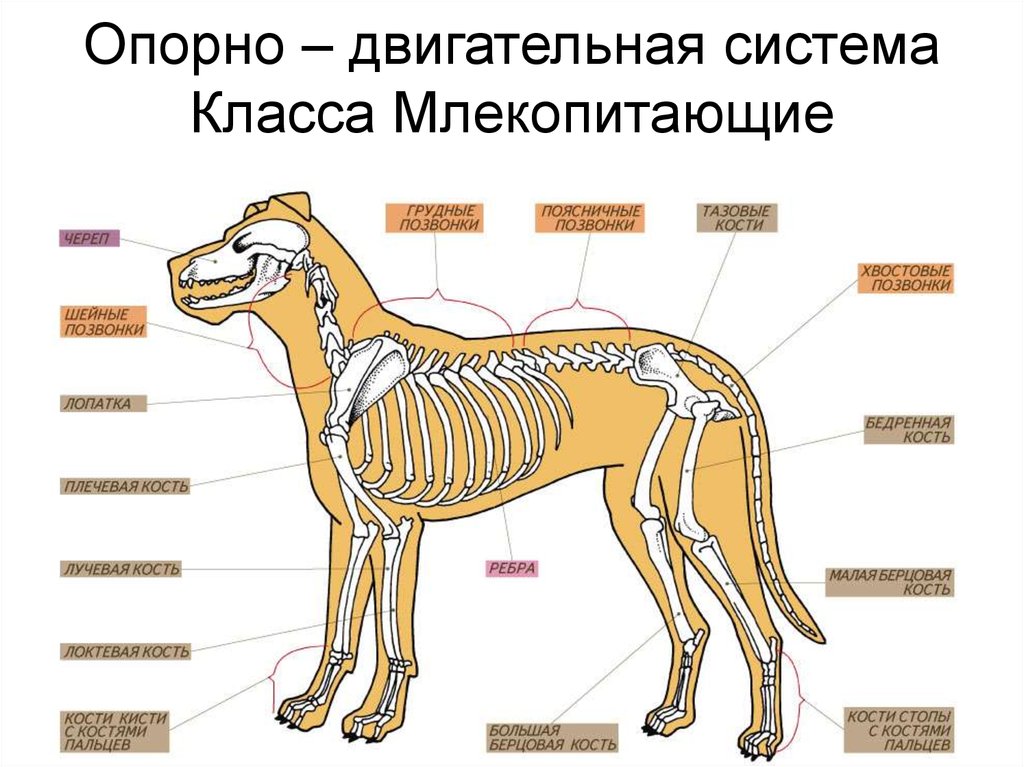
Нагромождение деревьев в лесу мешает видеть только крупным животным, таким как приматы. Более мелкие, например, белки, не испытывают таких затруднений, поскольку их небольшая голова может легко протиснуться между ветвями и листьями. Крупным животным, которые живут не в лесу, тоже вполне достаточно глаз, которые расположены по сторонам.
Автор фото, Thinkstock
Подпись к фото,Расположенные впереди глаза позволяли нашим предкам, жившим в лесу, видеть сквозь плотную листву и тесное переплетение веток
Таким образом, причина того, что глаза у нас находятся спереди, еще не установлена. У каждой гипотезы есть свои сильные и слабые стороны. Но независимо от того, зачем нам потребовалось такое зрение — чтобы прыгать с ветки на ветку, ловить вкусных жучков или видеть сквозь листву — очевидно, что такое расположение глаз связано с жизнью среди деревьев.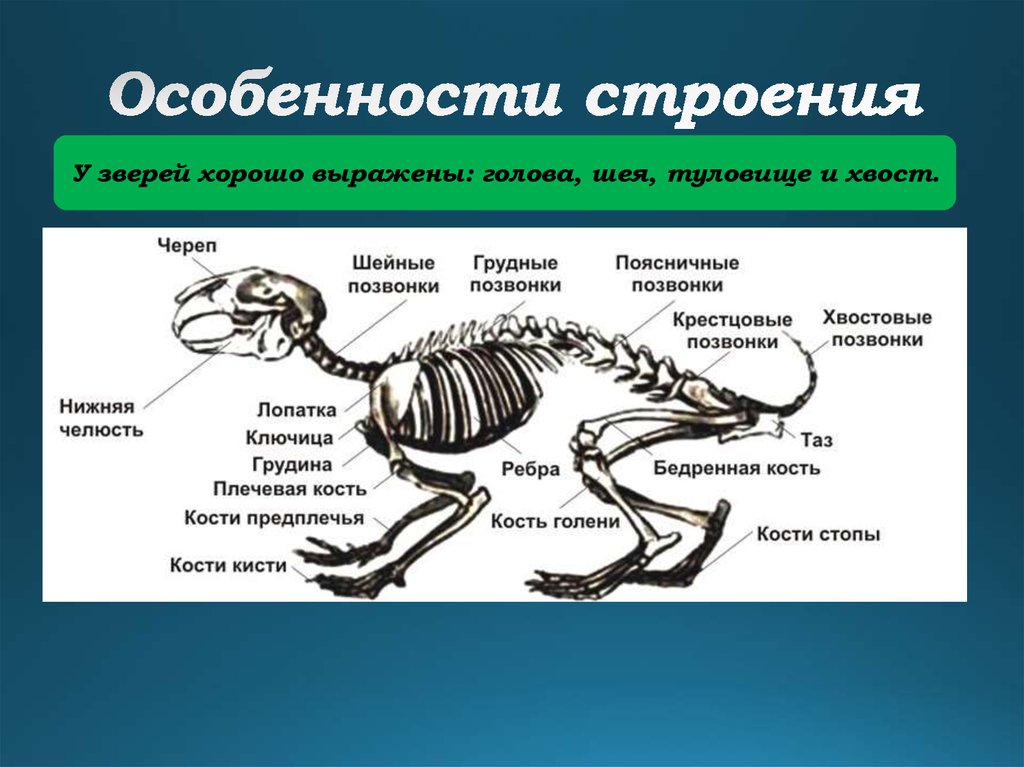
Прочитать оригинал этой статьи на английском языке можно на сайте BBC Future.
Все цвета, игнорируемые нашими глазами / Хабр
Большая часть млекопитающих полагается на обоняние больше, чем на зрение. Посмотрите на собачьи глаза – они расположены по бокам морды, не так, как у людей, у которых они находятся близко и направлены вперёд. Глаза по бокам позволяют увеличить область обзора, но плохо передают ощущению глубины и расстояние до объектов. Вместо хорошего зрения у собак, лошадей, мышей, антилоп – и в принципе у большинства млекопитающих – есть длинные влажные носы. Отличаемся от них мы, люди, человекообразные и обычные обезьяны. И у нашего зрения есть определённая необычная особенность, которую необходимо объяснить.
Со временем, занимая более освещённые экологические ниши, мы стали всё меньше полагаться на запах и всё больше на зрение. Мы потеряли влажные носы и рыльца, наши глаза подвинулись вперёд на лице и сблизились друг с другом, что улучшило наше умение оценивать расстояние (мы выработали улучшенное бинокулярное зрение).
Цветовое зрение работает через захват света с разными длинами волн и сравнение их с целью определения длин волн, отражённых объектом (то есть, его цвет). Синий цвет сильнее стимулирует рецептор, воспринимающий короткие длины волн, и слабо стимулирует рецептор, воспринимающий большие длины волн; красный цвет производит обратный эффект. Сравнивая относительную стимуляцию этих рецепторов, мы способны различать цвета.

Чтобы наилучшим образом воспринимать свет разных длин волн, колбочки должны равномерно располагаться по всему воспринимаемому людьми спектру, от 400 до 700 нм. Если мы посмотрим на распределение колбочек у шмеля, также обладающего трихроматическим зрением, мы также увидим равномерное распределение. И датчики цифровых камер тоже должны быть правильно расположены, чтобы правильно воспринимать цвета. Равномерное распределение колбочек/сенсоров обеспечивает хорошее спектральное прекрасное хроматическое покрытие для доступных длин волн. Но наше зрение работает не совсем так.
У нашего зрения нет такого равномерного спектрального распределения. У людей и прочих catarrhini области действия красных и зелёных колбочек пересекаются. Это означает, что мы ставим приоритет очень хорошее распознавание нескольких типов цветов – конкретно, красного и зелёного – за счёт невозможности увидеть так много цветов, как могли бы. Это странно. Почему же для нас так важно отличать красный от зелёного?
Тому было предложено несколько объяснений. Возможно, простейшее из них следующее: этот эффект является примером того, что биологи называют эволюционным ограничением. Ген, кодирующий рецептор зелёного цвета, и ген, кодирующий рецептор красного, появились в результате дупликации генов. Вероятно, что они изначально были почти одинаковыми по чувствительности, и времени на эволюционный отбор, в результате которого они стали бы разными, не хватило.
Возможно, простейшее из них следующее: этот эффект является примером того, что биологи называют эволюционным ограничением. Ген, кодирующий рецептор зелёного цвета, и ген, кодирующий рецептор красного, появились в результате дупликации генов. Вероятно, что они изначально были почти одинаковыми по чувствительности, и времени на эволюционный отбор, в результате которого они стали бы разными, не хватило.
Ещё одно объяснение подчёркивает эволюционные преимущества близкого соседства красных и зелёных колбочек. Поскольку оно позволяет нам хорошо различать зелёные и красные цвета, а также разбираться в разных оттенках розового и красного, у нас хорошо получается отличать зрелые фрукты, которые обычно меняются от зелёного к красному или оранжевому цветам при созревании. Свидетельств реальности этого эффекта найдено предостаточно. Трихроматическим людям гораздо лучше удаётся высматривать зрелые фрукты в зелёной листве, чем дихроматическим (которых обычно называют людьми с красно-зелёным дальтонизмом).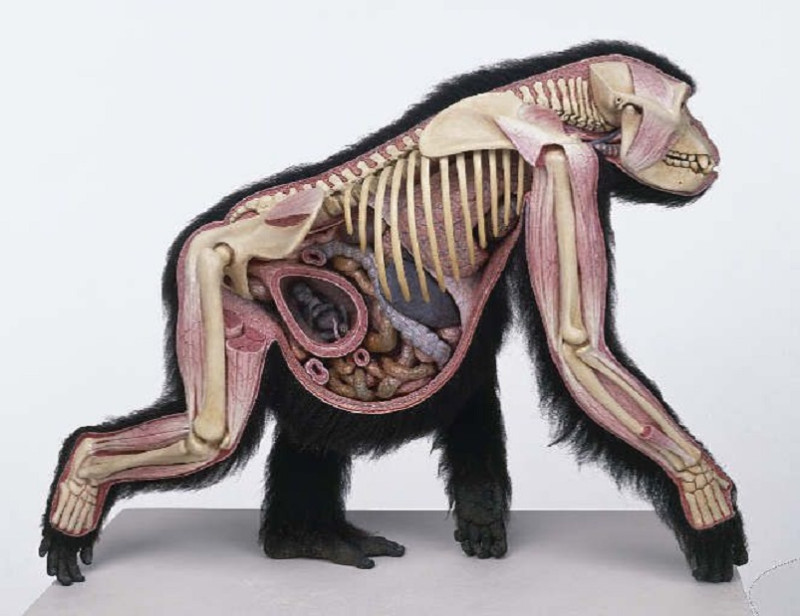 Что более важно, у нормально трихроматических людей это получается лучше, чем у людей, которым для эксперимента симулируют равномерно распределённый трихроматизм. У обезьян Нового света, среди которых некоторые трихроматические, а некоторые – дихроматические, первые распознают созревшие фрукты гораздо быстрее вторых, не пользуясь обонянием так сильно. Поскольку фрукты – критически важная часть диеты многих приматов, распознавание фруктов является правдоподобным фактором отбора, и не только для эволюции трихроматизма в общем, но и для нашей особенной, необычной формы трихроматизма.
Что более важно, у нормально трихроматических людей это получается лучше, чем у людей, которым для эксперимента симулируют равномерно распределённый трихроматизм. У обезьян Нового света, среди которых некоторые трихроматические, а некоторые – дихроматические, первые распознают созревшие фрукты гораздо быстрее вторых, не пользуясь обонянием так сильно. Поскольку фрукты – критически важная часть диеты многих приматов, распознавание фруктов является правдоподобным фактором отбора, и не только для эволюции трихроматизма в общем, но и для нашей особенной, необычной формы трихроматизма.
Итоговое объяснение связано с системой социальных сигналов. Многие виды приматов используют красный цвет, например, ярко-красный нос мандрила или красные пятна на груди гелады, при социальном общении. Точно так же эмоции людей сопровождаются изменением цвета лица, связанным с током крови, бледнея при недомогании или волнении, краснея при смущении и так далее. Возможно, распознавание таких признаков и сигналов может быть связанным с необычным распределением колбочек?
Недавно мы с коллегами проверили эту гипотезу экспериментально. Мы брали изображения морд самок макак-резусов, которые краснеют, когда самок интересует спаривание. Мы подготовили эксперименты, в которых люди смотрели на пары изображений одной и той же самки, на одном из которых её интересовало спаривание, а на другом – нет. Участников просили выбрать морду самки, интересующейся спариванием, но при этом мы немного подредактировали изображения. В некоторых подходах люди видели оригинальные изображения, в других они видели изображения с изменёнными цветами, эмулировавшими то, что увидел бы наблюдатель с другой системой восприятия цвета.
Мы брали изображения морд самок макак-резусов, которые краснеют, когда самок интересует спаривание. Мы подготовили эксперименты, в которых люди смотрели на пары изображений одной и той же самки, на одном из которых её интересовало спаривание, а на другом – нет. Участников просили выбрать морду самки, интересующейся спариванием, но при этом мы немного подредактировали изображения. В некоторых подходах люди видели оригинальные изображения, в других они видели изображения с изменёнными цветами, эмулировавшими то, что увидел бы наблюдатель с другой системой восприятия цвета.
Сравнивая таким способом разные виды трихроматизма и дихроматизма, мы обнаружили, что люди лучше всего справлялись с этой задачей, когда использовали нормальное человеческое трихроматическое зрение – и они гораздо лучше справлялись с задачей при помощи нормального зрения, чем с трихроматизмом с равномерным распределением колбочек (без наложения красного и зелёного спектров). Наши результаты совпали с гипотезой социальных сигналов: визуальная система людей лучше других справляется с обнаружением социальной информации на лицах других приматов.
Однако, мы проверили лишь необходимое условие гипотезы – то, что наше цветное зрение лучше справляется с этой задачей, чем другие возможные виды зрения. Возможно, что это сигналы появились в результате эволюции для того, чтобы воспользоваться чувствительностью наших глаз к определённым длинам волн, а не наоборот. Также возможно, что тут необходимо задействовать несколько объяснений одновременно. Один или несколько факторов могут оказаться связанными с происхождением распределения колбочек (к примеру, поедание фруктов), а другие факторы могут быть связанными с эволюционной поддержкой этого распределения после его появления в результате эволюции (к примеру, распознавание социальных сигналов).
Всё ещё точно неизвестно, почему у людей развилось такое странное цветное зрение. Возможно, это связано с добычей еды, социальными сигналами, эволюционными ограничениями, или какими-то другими объяснениями. Однако для исследования этого вопроса у нас есть много инструментов – генетическое секвенирование цветного зрения отдельного индивида, экспериментальная симуляция различных типов цветного зрения вкупе с поведенческим тестированием, наблюдения за дикими приматами, распознающими разные цвета.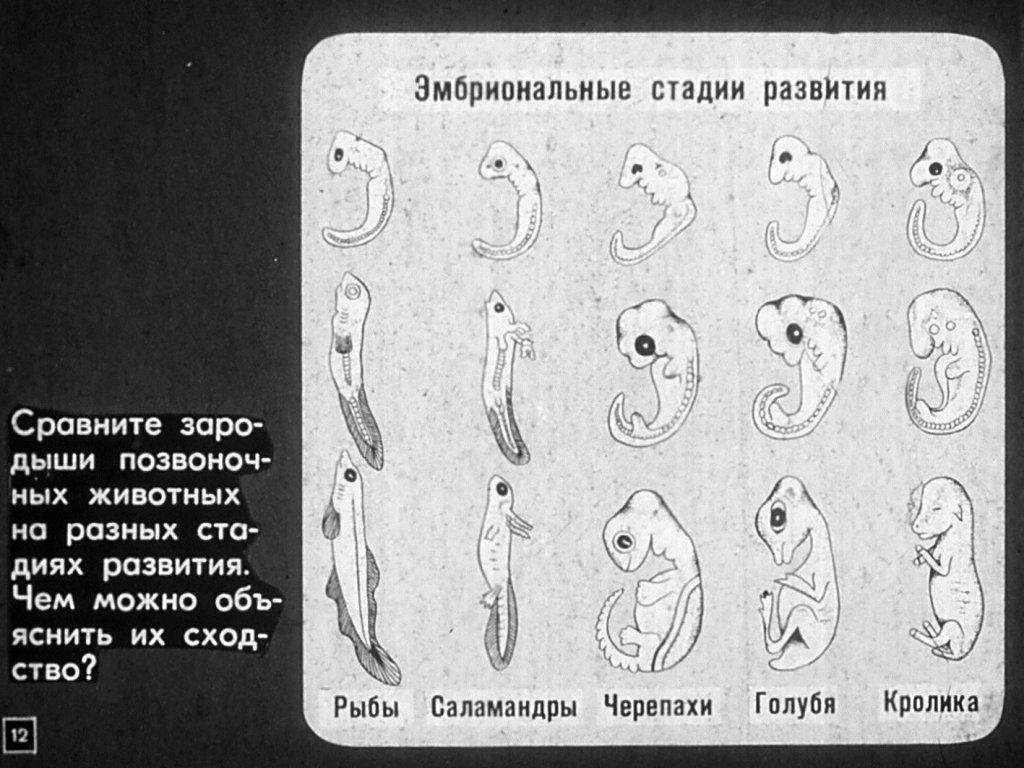 Есть что-то странное в том, как мы воспринимаем цвета. Мы поставили в приоритет способность различать несколько определённых цветов за счёт возможностей видеть столько цветов, сколько мы могли бы. Надеемся однажды узнать, почему так получилось.
Есть что-то странное в том, как мы воспринимаем цвета. Мы поставили в приоритет способность различать несколько определённых цветов за счёт возможностей видеть столько цветов, сколько мы могли бы. Надеемся однажды узнать, почему так получилось.
Обоняние и цветное зрение в эволюции млекопитающих развивались в противофазе • Александр Марков • Новости науки на «Элементах» • Генетика, Эволюция
Млекопитающие лишились цветного зрения в самом начале своей эволюции, потеряв два из четырех генов светочувствительных белков — опсинов. Впоследствии цветное зрение вновь появилось у обезьян благодаря дупликации одного из двух оставшихся опсинов. Как выяснилось, параллельно с утратой генов цветного зрения у древних млекопитающих в результате многочисленных дупликаций резко возросло количество генов обонятельных рецепторов. По-видимому, оба процесса — ослабление цветного зрения и развитие обоняния — были связаны с тем, что экспансия динозавров в конце триасового — начале юрского периода вынудила древних млекопитающих перейти к ночному образу жизни.
Эволюция цветного зрения у позвоночных изучена достаточно подробно. Способность различать цвета определяется светочувствительными белками колбочек — опсинами, которые могут быть «настроены» на разную длину волны. В зависимости от того, какие аминокислоты стоят в определенных «ключевых» позициях в молекуле опсина, белок избирательно реагирует на световые волны той или иной длины. Еще до выхода на сушу позвоночные выработали весьма совершенную систему цветного зрения, основанную на четырех опсинах (тетрахроматическое зрение). Эта система сохранилась у многих наземных позвоночных, включая птиц, которые великолепно различают цвета. Возможно, если бы такое зрение имелось и у людей, нам казалась бы убогой трихроматическая система отображения цвета, используемая в наших телевизорах и компьютерных мониторах. У человека, как и у всех обезьян Старого Света, зрение трихроматическое. У большинства других млекопитающих из четырех опсинов, имевшихся у древних позвоночных, сохранилось только два (дихроматическое зрение). Предки обезьян тоже имели дихроматическое зрение (а значит, не могли отличить красный цвет от зеленого).
Предки обезьян тоже имели дихроматическое зрение (а значит, не могли отличить красный цвет от зеленого).
Считается, что утрата млекопитающими двух опсинов была связана с тем, что их предки когда-то перешли к ночному образу жизни. Скорее всего, это произошло еще на заре их истории — в конце триасового или в начале юрского периода — и было связано с перипетиями долгой конкурентной борьбы между двумя основными эволюционными стволами наземных позвоночных — синапсидами и диапсидами. В пермском периоде синапсидные рептилии — предки млекопитающих — были господствующей группой. В следующем, триасовом периоде их господство пошатнулось, поскольку на сцене появились молодые активные конкуренты — архозавры, относящиеся к группе диапсидных рептилий. Архозавры делали ставку на крупные размеры, быстрый бег и острые зубы, а у синапсидных рептилий тем временем бурно шла «маммализация» — развитие черт млекопитающих. Они понемногу мельчали и уходили «в тень».
В конце триаса — начале юры власть на суше окончательно перешла к одной из групп архозавров, а именно к динозаврам. Синапсиды вымерли почти полностью, за исключением одной небольшой группы, которая дала начало млекопитающим. В течение всего юрского и мелового периодов, до самого вымирания динозавров, млекопитающим приходилось вести преимущественно ночной образ жизни, и к тому же оставаться маленькими, чтобы пореже попадаться на глаза господствующим дневным хищникам. В этих обстоятельствах цветное зрение стало бесполезным, и два опсиновых гена были потеряны. Естественный отбор не умеет заглядывать в будущее — он сохраняет только те признаки и гены, которые нужны здесь и сейчас. Когда после вымирания динозавров многие млекопитающие снова стали дневными, им пришлось обходиться дихроматическим зрением, поскольку взять новые опсиновые гены взамен утраченных было негде.
Синапсиды вымерли почти полностью, за исключением одной небольшой группы, которая дала начало млекопитающим. В течение всего юрского и мелового периодов, до самого вымирания динозавров, млекопитающим приходилось вести преимущественно ночной образ жизни, и к тому же оставаться маленькими, чтобы пореже попадаться на глаза господствующим дневным хищникам. В этих обстоятельствах цветное зрение стало бесполезным, и два опсиновых гена были потеряны. Естественный отбор не умеет заглядывать в будущее — он сохраняет только те признаки и гены, которые нужны здесь и сейчас. Когда после вымирания динозавров многие млекопитающие снова стали дневными, им пришлось обходиться дихроматическим зрением, поскольку взять новые опсиновые гены взамен утраченных было негде.
До недавних пор ученые предполагали, что оба опсиновых гена были утрачены практически сразу и очень давно, еще до разделения млекопитающих на однопроходных и териевых (= сумчатые + плацентарные). Однако один из потерянных генов обнаружился в геноме утконоса. Это значит, что гены были потеряны не сразу, а по очереди, и не так быстро. Общий предок всех современных млекопитающих еще имел три опсина, а общий предок териевых — уже только два. Некоторые австралийские сумчатые вроде бы имеют полноценное цветное зрение, но ни одного из двух потерянных генов в их геномах обнаружить не удалось, несмотря на целенаправленные поиски. Значит, если у них действительно есть цветное зрение, оно приобретено ими вторично и на иной генетической основе.
Это значит, что гены были потеряны не сразу, а по очереди, и не так быстро. Общий предок всех современных млекопитающих еще имел три опсина, а общий предок териевых — уже только два. Некоторые австралийские сумчатые вроде бы имеют полноценное цветное зрение, но ни одного из двух потерянных генов в их геномах обнаружить не удалось, несмотря на целенаправленные поиски. Значит, если у них действительно есть цветное зрение, оно приобретено ими вторично и на иной генетической основе.
Как это в принципе может происходить, отлично показывает пример обезьян. У общего предка обезьян Старого Света, который жил 30–40 млн лет назад, один из двух сохранившихся опсиновых генов подвергся дупликации, и естественный отбор быстро «настроил» получившиеся копии на разные длины волн. Для этого потребовалось зафиксировать всего-навсего три мутации (аминокислотные замены). В итоге зрение у обезьян стало трихроматическим, что дало им возможность отличать спелые плоды от зеленых и свежую листву (наиболее питательную) от старой (у многих тропических растений молодые листья имеют красноватый оттенок). Параллельно и совершенно независимо сформировалось цветное зрение у некоторых обезьян Нового Света. У них недостающий третий опсин возник как аллельный вариант одного из двух старых опсиновых генов. Этот опсиновый ген расположен в X-хромосоме, поэтому шанс получить от родителей три разных опсина (и трихроматическое зрение) есть только у самок, и то не у всех. Но обезьяны ведут общественный образ жизни, и наличие в стаде хотя бы нескольких самок, способных отличить красное от зеленого, оказывается очень полезным для всего коллектива.
Параллельно и совершенно независимо сформировалось цветное зрение у некоторых обезьян Нового Света. У них недостающий третий опсин возник как аллельный вариант одного из двух старых опсиновых генов. Этот опсиновый ген расположен в X-хромосоме, поэтому шанс получить от родителей три разных опсина (и трихроматическое зрение) есть только у самок, и то не у всех. Но обезьяны ведут общественный образ жизни, и наличие в стаде хотя бы нескольких самок, способных отличить красное от зеленого, оказывается очень полезным для всего коллектива.
Такуси Кисида (Takushi Kishida) из Киотского университета в своей статье, опубликованной в журнале PLoS ONE, показал, что гены обонятельных рецепторов наземных позвоночных имеют не менее увлекательную эволюционную историю, чем гены зрительных белков. Как выяснилось, уменьшение числа опсинов сопровождалось увеличением числа обонятельных рецепторов, и наоборот.
Млекопитающие отличаются от других наземных позвоночных не только проблемами с цветным зрением, но и гораздо более развитым обонянием. Например, у крысы насчитывается до 1600 функционирующих генов обонятельных рецепторов, тогда как у курицы их всего около 80. В отличие от цветов спектра, для различения многочисленных запахов тремя-четырьмя генами не обойтись: на каждую летучую молекулу нужен свой рецептор. Исследователи давно установили, что многочисленные гены обонятельных рецепторов млекопитающих возникли в результате множественных дупликаций из исходного небольшого набора. Естественно было предположить, что развитие обоняния у млекопитающих, как и утрата цветного зрения, было связано с переходом к ночному образу жизни. В этом случае большинство дупликаций обонятельных генов должно было произойти примерно в то же время, что и утрата двух опсиновых генов.
Например, у крысы насчитывается до 1600 функционирующих генов обонятельных рецепторов, тогда как у курицы их всего около 80. В отличие от цветов спектра, для различения многочисленных запахов тремя-четырьмя генами не обойтись: на каждую летучую молекулу нужен свой рецептор. Исследователи давно установили, что многочисленные гены обонятельных рецепторов млекопитающих возникли в результате множественных дупликаций из исходного небольшого набора. Естественно было предположить, что развитие обоняния у млекопитающих, как и утрата цветного зрения, было связано с переходом к ночному образу жизни. В этом случае большинство дупликаций обонятельных генов должно было произойти примерно в то же время, что и утрата двух опсиновых генов.
Чтобы проверить эту гипотезу, Кисида провел тщательный сравнительный анализ генов обонятельных рецепторов в геномах шести наземных позвоночных: лягушки, курицы, утконоса, опоссума, собаки и мыши. Анализ этих шести видов позволяет реконструировать ситуацию в важнейших точках ветвления эволюционного древа наземных позвоночных.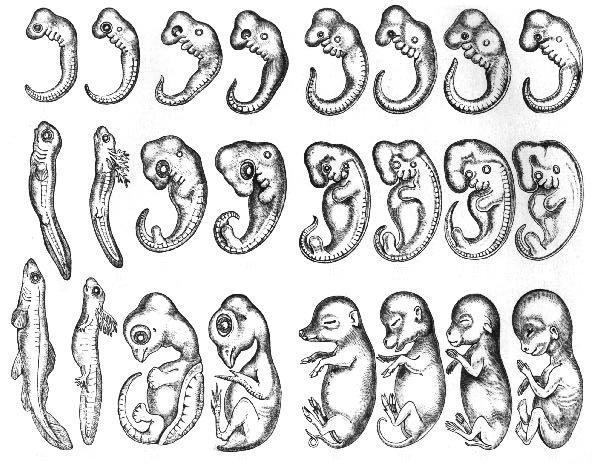 Сравнение лягушки с другими видами проливает свет на общего предка современных амфибий и амниот (= рептилии + птицы + млекопитающие), жившего около 340 млн лет назад (начало каменноугольного периода). Курица, как прямой потомок архозавров, помогает составить представление об общем предке синапсидных и диапсидных тетрапод, жившем около 310 млн лет назад (вторая половина каменноугольного периода). Утконос расскажет об общем предке однопроходных и териевых (180 млн лет назад, ранняя юра), опоссум — об общем предке сумчатых и плацентарных (140 млн лет назад, ранний мел). Эволюционные пути предков собаки и мыши разошлись около 85 млн лет назад (поздний мел). Что касается нас с вами, то мы на этом упрощенном эволюционном древе ближе всего к мыши.
Сравнение лягушки с другими видами проливает свет на общего предка современных амфибий и амниот (= рептилии + птицы + млекопитающие), жившего около 340 млн лет назад (начало каменноугольного периода). Курица, как прямой потомок архозавров, помогает составить представление об общем предке синапсидных и диапсидных тетрапод, жившем около 310 млн лет назад (вторая половина каменноугольного периода). Утконос расскажет об общем предке однопроходных и териевых (180 млн лет назад, ранняя юра), опоссум — об общем предке сумчатых и плацентарных (140 млн лет назад, ранний мел). Эволюционные пути предков собаки и мыши разошлись около 85 млн лет назад (поздний мел). Что касается нас с вами, то мы на этом упрощенном эволюционном древе ближе всего к мыши.
Результаты, полученные Кисидой, говорят о том, что у общего предка амфибий и амниот было примерно 100–110 генов обонятельных рецепторов. Изначально большинство этих генов находилось на одной хромосоме, но время от времени они перепрыгивали на другие хромосомы. Этот процесс «рассеивания» обонятельных генов по хромосомам, по-видимому, шел уже у первых наземных позвоночных и практически прекратился к моменту расхождения линий однопроходных и териевых (180 млн лет назад). В результате у всех млекопитающих обонятельные гены присутствуют почти на всех хромосомах. У человека, например, их нет только на двух хромосомах: 20-й и Y-хромосоме. Кисида предполагает, что рассеивание обонятельных генов по хромосомам облегчило их последующую множественную дупликацию.
Этот процесс «рассеивания» обонятельных генов по хромосомам, по-видимому, шел уже у первых наземных позвоночных и практически прекратился к моменту расхождения линий однопроходных и териевых (180 млн лет назад). В результате у всех млекопитающих обонятельные гены присутствуют почти на всех хромосомах. У человека, например, их нет только на двух хромосомах: 20-й и Y-хромосоме. Кисида предполагает, что рассеивание обонятельных генов по хромосомам облегчило их последующую множественную дупликацию.
У общего предка синапсидных и диапсидных число обонятельных генов осталось прежним (около сотни). У общего предка однопроходных и териевых их было уже около 330, у общего предка плацентарных и сумчатых их число возросло примерно до 670. Общий предок мышей и собак имел около 740 генов обонятельных рецепторов.
Самый главный результат, полученный Кисидой, состоит в том, что практически все дупликации обонятельных генов в эволюции тетрапод были приурочены к отрезку эволюционного древа, заключенному между общими предками диапсидных и синапсидных (310 млн лет назад) и плацентарных и сумчатых (140 млн лет назад). В целом дупликации, по сравнению с «рассеиванием», позже начались и позже закончились.
В целом дупликации, по сравнению с «рассеиванием», позже начались и позже закончились.
Учитывая разрешающую способность примененных методик, можно заключить, что в пределах этой разрешающей способности период массовых дупликаций обонятельных генов в точности совпадает с периодом утраты опсиновых генов. Первый опсиновый ген был потерян на отрезке между общим предком диапсидных и синапсидных и общим предком однопроходных и териевых, то есть на ранних этапах становления млекопитающих. Второй опсиновый ген был потерян на отрезке между общим предком однопроходных и териевых и общим предком сумчатых и плацентарных, то есть на ранних этапах эволюции териевых. Именно к этим двум отрезкам приурочены и дупликации обонятельных генов. Если бы существовало какое-нибудь современное животное, предки которого отделились бы от «нашего» эволюционного ствола позже курицы, но раньше утконоса, датировки можно было бы существенно уточнить. Но такого животного, к сожалению, нет.
Но такого животного, к сожалению, нет.
Кисида также отмечает, что восстановление цветного зрения у обезьян Старого Света сопровождалось утратой значительной части обонятельных генов (или превращением их в неработающие псевдогены). Очевидно, развитие зрения и обоняния происходило в противофазе. Когда древние млекопитающие перешли к ночному образу жизни, роль зрения уменьшилась, а роль обоняния возросла. Когда обезьяны вернулись к дневной жизни и стали снова полагаться в основном на зрение, их обоняние ослабло.
Еще одна любопытная деталь состоит в том, что одно из семейств обонятельных генов (известное как семейство №7; у приматов это самое многочисленное семейство обонятельных генов) подвергалось усиленной дупликации уже после разделения плацентарных и сумчатых. О некоторых рецепторах этого семейства известно, что они реагируют на половые феромоны.
P.S. У читателей может возникнуть резонный вопрос: неужели для улучшения зрения или обоняния достаточно добавить новый рецептор? А откуда возьмутся новые мозговые структуры, которые должны обрабатывать сигналы от этого нового рецептора? Похоже на то, что новых мозговых структур для этого не требуется — имеющиеся структуры используют какие-то обобщенные, универсальные алгоритмы интерпретации поступающих сигналов. По-видимому, мозг в процессе индивидуального развития автоматически обучается различать сигналы, приходящие от разных рецепторов, и интерпретировать их именно как разные сигналы. Ничего не меняя в структуре мозга, можно добавить в сетчатку новый опсин, и мозг сам разберется, что делать с новым типом сигналов. См. об этом: Мышиный мозг готов увидеть мир по-человечески, «Элементы», 29.03.2007, Восприятием цвета заведует мозг, «Элементы», 28.10.2005.
По-видимому, мозг в процессе индивидуального развития автоматически обучается различать сигналы, приходящие от разных рецепторов, и интерпретировать их именно как разные сигналы. Ничего не меняя в структуре мозга, можно добавить в сетчатку новый опсин, и мозг сам разберется, что делать с новым типом сигналов. См. об этом: Мышиный мозг готов увидеть мир по-человечески, «Элементы», 29.03.2007, Восприятием цвета заведует мозг, «Элементы», 28.10.2005.
Источник: Takushi Kishida. Pattern of the Divergence of Olfactory Receptor Genes during Tetrapod Evolution // PLoS ONE. 2008. V. 3. P. e2385.
Об эволюционной роли дупликации генов см. также:
Прослежена эволюционная история одного из человеческих генов, «Элементы», 17.06.2008.
Александр Марков
Особенности механизмов зрения птиц
Статья на конкурс «Био/Мол/Текст»: Птицы — удивительные создания, господствующие в воздухе среди всех хордовых. Численность видов птиц на три тысячи превосходит млекопитающих, а их оперение никогда не тускнеет. Имея физиологию, отличную от млекопитающих, они получили заметные эволюционные преимущества. Одно из них — уникальное зрение, совмещающее в себе навигацию по магнитному полю, невероятную остроту и восприятие ультрафиолета.
Имея физиологию, отличную от млекопитающих, они получили заметные эволюционные преимущества. Одно из них — уникальное зрение, совмещающее в себе навигацию по магнитному полю, невероятную остроту и восприятие ультрафиолета.
Эта работа опубликована в номинации «Свободная тема» конкурса «Био/Мол/Текст»-2021/2022.
Партнер номинации — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Генеральный партнер конкурса — международная инновационная биотехнологическая компания BIOCAD.
Генеральный партнер конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Словарик
- Колбочки
- один из двух типов фоторецепторов сетчатки глаза. Чувствительны к свету благодаря наличию молекул йодопсина. Отвечают за цветовое зрение.
- Палочки
- второй тип фоторецепторов.
 Чувствительны к свету низкой яркости благодаря наличию молекул родопсина.
Чувствительны к свету низкой яркости благодаря наличию молекул родопсина. - Ретиналь
- каротиноид, связанный с белками-опсинами. Является химической основой зрения позвоночных животных. Вместе с опсином палочек образует комплекс — родопсин. В колбочках образует комплексы йодопсин, эритролаб и хлоролаб. Разница цветовосприятия светочувствительных комплексов связана с аминокислотной последовательностью боковых цепей опсина.
- Родопсин
- основной зрительный пигмент палочек сетчатки глаза. Рекомендуем почитать статью «Биомолекулы» «Зрительный родопсин — рецептор, реагирующий на свет» [1].
- Фотоизомеризация
- переход молекулы из одного изомерного состояния (конформации) в другое под действием света.
«Видимый» диапазон света. Так ли было всегда?
Физиология зрения и физика света тесно друг с другом связаны. В 1927 году академик С.И. Вавилов заканчивает свою книгу «Глаз и Солнце» словами: «Глаз нельзя понять, не зная Солнца. Вот почему глаз солнечен, по словам поэта» [2].
Вот почему глаз солнечен, по словам поэта» [2].
В чем же «солнечность» глаза? Для того, чтобы понять это, достаточно взглянуть на электромагнитный спектр излучения Солнца (рис. 1, сверху). Именно благодаря спектру испускания Солнца сформировались спектральные характеристики светочувствительных зрительных пигментов — палочек и колбочек, запускающих процесс зрения. На самом деле, человеческий глаз способен различить не весь спектральный диапазон, доходящий до поверхности Земли (рис. 1, снизу), но именно та его часть, которую могут распознать наши зрительные пигменты, названа «видимым светом» — 380–760 нм [3].
Рисунок 1. Электромагнитный спектр солнечного излучения. У поверхности Земли около 80% солнечной энергии ограничено спектральным диапазоном 300–1100 нм [3]. При этом «видимым светом» принято обозначать только ту область, которую способен распознавать человеческий глаз.
«Википедия»
В ходе биологической эволюции зрительная клетка достигла предела чувствительности, допускаемой квантовой теорией света. Согласно данным физиков из Университета Иллинойса, человеку достаточно всего трех квантов света, чтобы увидеть вспышку, а отдельные клетки сетчатки (палочки) могут регистрировать и единичные кванты света. Однако вероятность, что сигнал, порождаемый только одной клеткой, дойдет до мозга, крайне мала [3]. Получается цепочка: один квант → поглотившая его молекула зрительного пигмента → одна возбужденная клетка сетчатки (палочка) → зрительный сигнал. Но чтобы распознать еле заметную вспышку света, одного кванта недостаточно. Обычно человеку требуется около 10–20 квантов света; тогда сигналы от 10–20 палочек суммируются и передаются в мозг, в его зрительные центры. Благодаря этому механизму человек способен различать на ночном небе очень слабые звездочки — шестой, а в некоторых случаях даже седьмой и восьмой звездной величины. Доходящий от звезд до Земли свет — это всего лишь 10−12—10−14 от полного солнечного света. Таким образом, наша зрительная система (глаза и мозг) может эффективно работать в огромном диапазоне освещенностей — от почти полной темноты до яркого солнечного дня.
Согласно данным физиков из Университета Иллинойса, человеку достаточно всего трех квантов света, чтобы увидеть вспышку, а отдельные клетки сетчатки (палочки) могут регистрировать и единичные кванты света. Однако вероятность, что сигнал, порождаемый только одной клеткой, дойдет до мозга, крайне мала [3]. Получается цепочка: один квант → поглотившая его молекула зрительного пигмента → одна возбужденная клетка сетчатки (палочка) → зрительный сигнал. Но чтобы распознать еле заметную вспышку света, одного кванта недостаточно. Обычно человеку требуется около 10–20 квантов света; тогда сигналы от 10–20 палочек суммируются и передаются в мозг, в его зрительные центры. Благодаря этому механизму человек способен различать на ночном небе очень слабые звездочки — шестой, а в некоторых случаях даже седьмой и восьмой звездной величины. Доходящий от звезд до Земли свет — это всего лишь 10−12—10−14 от полного солнечного света. Таким образом, наша зрительная система (глаза и мозг) может эффективно работать в огромном диапазоне освещенностей — от почти полной темноты до яркого солнечного дня.
Глаз животных и человека как нельзя лучше приспособлен для восприятия именно естественного солнечного света, который и сформировал «под себя» органы зрения всех животных — от самых примитивных беспозвоночных до высших позвоночных.
Фотоэлектронный умножитель глаза
Но как возможно, чтобы всего один квант света вызывал бурную реакцию в нашем глазу? «Биология становится слишком серьезной наукой, чтобы ее можно было доверять биологам», — пошутил кто-то из физиков. И оказался абсолютно прав!
Чтобы понять, как работает колбочка, надо разобраться в устройстве такого прибора, как фотоэлектронный умножитель (ФЭУ). Принципиальная схема его работы представлена на рисунке 3А (см. также врезку) и заключается в каскадном последовательном выбивании электронов с фотопластинок для увеличения количества электронов, попадающих на детектор. В случае с глазом, в роли детектора выступает нервное окончание.
Маленький прибор с большими перспективами для человечества
ФЭУ представляет собой электровакуумный прибор, в котором поток электронов, попадая на специальные пластинки, усиливается за счет вторичной эмиссии — то есть происходит повторное выбивание электронов из пластинки, обозначенной на схеме как динод (рис. 2А). Так, получается, что при попадании на первую пластинку динода всего одного электрона на выходе к коллектору (аноду) будет уже поток электронов, превышающий в 105 раз (и выше!) исходный сигнал. ФЭУ, созданные советским изобретателем Л.А. Кубецким (рис. 2В) в 1930–1934 годы, широко используются в телевизорах, приборах ночного видения и прочей привычной нам технике. Также ФЭУ нашли огромное применение в приборах современной оптики и микроскопии, без которых было бы невозможно разглядеть клетки, из которых состоит все живое [4–6]!
2А). Так, получается, что при попадании на первую пластинку динода всего одного электрона на выходе к коллектору (аноду) будет уже поток электронов, превышающий в 105 раз (и выше!) исходный сигнал. ФЭУ, созданные советским изобретателем Л.А. Кубецким (рис. 2В) в 1930–1934 годы, широко используются в телевизорах, приборах ночного видения и прочей привычной нам технике. Также ФЭУ нашли огромное применение в приборах современной оптики и микроскопии, без которых было бы невозможно разглядеть клетки, из которых состоит все живое [4–6]!
Рисунок 2А. Фотоумножители электронные и природные. Схема фотоэлектронного умножителя (ФЭУ).
портал САМ
Рисунок 2Б. Фотоумножители электронные и природные. Схематическое изображение колбочки глаза.
Рисунок 2В. Леонид Александрович Кубецкий. Человек, которого должны знать в лицо. Без его изобретений был бы невозможен просмотр телевизора!
Без его изобретений был бы невозможен просмотр телевизора!
«Википедия»
Однако количество электронов, которые возможно выбить из пластинки динода, не бесконечно. Поэтому когда ФЭУ приходят в негодность, их меняют. А что же с глазом? Если посмотреть на схему строения колбочки глаза (рис. 2Б), то мы увидим, что ее наружный сегмент состоит из плазматической мембраны, наполненной стопками дисков с молекулами светочувствительных белков — родопсинов. В палочке квант света поглощается одной из 109 молекул родопсина. В результате чрезвычайно быстрой (менее 1 пикосекунды — 10−12 с) фотохимической реакции и затем последовательности темновых реакций, зависимых от температуры превращений, молекула родопсина обесцвечивается. Это означает, что каким-то образом в палочке эта одна обесцвеченная из 109 необесцвеченных молекул должна быть «узнана». Далее эта «узнанная» молекула запускает мощный каскад усиления первичного светового, фотохимического события, вследствие чего клетка отвечает миниатюрным, хотя и вполне ощутимым электрическим рецепторным сигналом. В последние годы удалось напрямую зарегистрировать этот миниатюрный сигнал.
В последние годы удалось напрямую зарегистрировать этот миниатюрный сигнал.
Миниатюрный электрический ответ в палочке на один квант все же достаточно велик: он составляет около 3% от ее максимального ответа на яркую, насыщенную вспышку света. Такой величины сигнал может быть уверенно передан дальше — нейронам сетчатки, а затем в мозг.
Когда молекула родопсина «изживает» себя, диск за ненадобностью выкидывается из палочки, но так никуда и не исчезает из глаза. Именно поэтому у людей преклонного возраста белок глаза имеет желтоватый оттенок — это не что иное, как накопившиеся израсходованные диски с родопсином [3], [7].
Родопсин и фотоизомеризация. Молекула, благодаря которой мы видим свет
«Биомолекула» уже подробно рассказывала об этой замечательной молекуле, и прежде всего мы советуем читателям обратиться к статье «Зрительный родопсин — рецептор, реагирующий на свет» [8]. Являясь типичным представителем G-белок сопряженных рецепторов (подробнее о G-белках читайте в статье «Биомолекулы» «Аллостерические регуляторы GPCR: ключи от всех замков» [9]), родопсин переходит из неактивной в активную форму при поглощении кванта света ретиналем (рис. 3): главной реакцией этого фотопревращения является переход хромофорной группы родопсина — 11-cis-ретиналя — в all-trans-ретиналь.
3): главной реакцией этого фотопревращения является переход хромофорной группы родопсина — 11-cis-ретиналя — в all-trans-ретиналь.
Рисунок 3А. Схема фотопревращений родопсина. Промежуточные стадии фотопревращения родопсина при попадании кванта света. При этом конформационные перестройки белковой части молекулы происходят сначала в хромофорном центре, а затем и в цитоплазматическом домене. Родопсин переходит в активированное состояние и взаимодействует с G-белком трансдуцином (подробнее — в статье «Зрительный родопсин — рецептор, реагирующий на свет» [8]).
Рисунок 3Б. Молекула 11-cis- и all-trans-ретиналя.
Птичье превосходство
Зрение птиц отличается от человеческого и продиктовано их образом жизни. Обитание в воздушной среде, высокая маневренность и широкий обзор горизонта при наборе высоты способствовали максимальному развитию зрения у этого класса животных. Формированию детального изображения при высокой остроте способствуют крупные глаза, совершенная аккомодация хрусталика с помощью системы поперечнополосатых цилиарных мышц, возможность дополнительно изменить кривизну роговицы, васкуляризованный гребешок внутри глаза и сложно устроенная сетчатка с одной или двумя фовекальными ямками, не имеющая аналогов у других классов животных. До 1980-х годов изучению глаз птиц не уделяли особенного внимания, поэтому известно далеко не так много, как хотелось бы. Важный вклад в изучение глаз птиц внес Боумейкер с коллегами, изучив спектры поглощения зрительных пигментов фоторецепторов птиц [10]. Активное изучение механизмов зрения птиц началось в 2000-х годах. На сегодня известно, что обработку зрительного сигнала обеспечивают палочки и колбочки, но их свойства, морфология и биохимия отличны от человеческих и малоизучены. Механизм цветовосприятия у птиц на сегодняшний день так и не установлен.
Формированию детального изображения при высокой остроте способствуют крупные глаза, совершенная аккомодация хрусталика с помощью системы поперечнополосатых цилиарных мышц, возможность дополнительно изменить кривизну роговицы, васкуляризованный гребешок внутри глаза и сложно устроенная сетчатка с одной или двумя фовекальными ямками, не имеющая аналогов у других классов животных. До 1980-х годов изучению глаз птиц не уделяли особенного внимания, поэтому известно далеко не так много, как хотелось бы. Важный вклад в изучение глаз птиц внес Боумейкер с коллегами, изучив спектры поглощения зрительных пигментов фоторецепторов птиц [10]. Активное изучение механизмов зрения птиц началось в 2000-х годах. На сегодня известно, что обработку зрительного сигнала обеспечивают палочки и колбочки, но их свойства, морфология и биохимия отличны от человеческих и малоизучены. Механизм цветовосприятия у птиц на сегодняшний день так и не установлен.
Подробнее о том, как видят другие животные, можно прочитать в статьях «Биомолекулы» «Разные глаза — разные возможности» [11] и «Откуда пошло зрение» [12].
Вижу на все 360
o (кроме кончика клюва)Зрение — основное чувство дальней и ближней ориентации птиц. В отличие от других позвоночных, среди них нет ни одного вида с редуцированными или слепыми глазами. У птиц, ведущих ночной образ жизни, можно наблюдать трубчатые глаза с небольшим количеством детекторов цвета, но с многочисленными палочками, которые хорошо функционируют при слабом освещении. Аккомодация глаза достигается изменением формы хрусталика и его перемещением [13].
На рисунке 4 хорошо видно, что глаза птиц расположены таким образом, чтобы обеспечивать хорошее бинокулярное зрение. Различное расположение глаз (рис. 4Б, 4В) связано с особенностями жизни каждого вида. Так, сове, будучи ночным хищником, вовсе не обязательно видеть то, что творится у нее за спиной, в то время как цапля или вальдшнеп, постоянно пребывающие в поиске еды, имеют полный обзор вокруг своей головы. Все это сводит к минимуму возможность незаметно подкрасться к птице сзади. В то же время, если птица будет полностью сконцентрирована на изучении объекта вдалеке, она может не заметить подкрадывающегося хищника. Подробнее об этом мы поговорим в других разделах.
Подробнее об этом мы поговорим в других разделах.
Если взглянуть на рисунок 4Б, то видно, что у вальдшнепа есть слепая зона на самом затылке и кончике клюва. Аналогично — у ястреба, воробья и цапли. Вот так: получается, что несмотря на широкий угол зрения, птица почти никогда не видит, что расположено у нее прямо под клювом.
Рисунок 4А. Поле зрения у разных видов птиц и других животных. На этой панели показано поле зрения ястреба с указанием зон моно- и бинокулярного зрения. Продолжение в крутилке!
«Биология и медицина»
Рисунок 4Б. Сравнение зон монокулярного и бинокулярного зрения у птиц разных видов.
«Уроки биологии»
Рисунок 4В. Сравнение зон монокулярного и бинокулярного зрения у птиц разных видов.
адаптировано из Medical Illustration & Animation
Рисунок 4Г. Поля зрения у зайца, совы и человека. У хищников больше развито бинокулярное зрение.
Поля зрения у зайца, совы и человека. У хищников больше развито бинокулярное зрение.
GlazExpert
Как же они видят?
Глазное яблоко птиц — сложный рецепторный аппарат, основные структуры которого сходны со структурами глаз других позвоночных (рис. 6). Орган зрения состоит из глазного яблока со зрительным нервом и придаточных органов: век, слезных желез и двигательных мышц. Глазное яблоко спереди выпуклое, в задней части округлое, в середине в виде усеченного конуса. Яблоко имеет три оболочки — наружную, среднюю и внутреннюю. Сосудистая (средняя) оболочка находится под склерой; она состоит из собственно сосудистой оболочки, радужной оболочки и ресничного тела.
К центру сетчатки расположена центральная ямка с большой плотностью рецепторов. Это область наибольшей остроты переднего зрения, то есть резкого и четкого обнаружения объектов. У 54% птиц есть вторая центральная ямка для улучшенного бокового обзора. На оптическом диске у птиц есть небольшое слепое пятно, лишенное фоторецепторов; это место присоединения зрительного нерва и сосудов к глазу.
Птичий глаз напоминает глаз рептилии: он оснащен цилиарной мышцей, способной менять форму хрусталика быстрее и сильнее, чем у млекопитающих. В животном царстве у птиц самые большие глаза относительно размера тела. По внутреннему строению они схожи с глазами других позвоночных, но имеют уникальную структуру — гребень. Глаза птиц защищены третьей прозрачной подвижной мембраной.
Рисунок 5. Глаз ночной хищной птицы. Глазное яблоко птиц имеет уплощенную форму и малоподвижно. Однако этот «недостаток» природа компенсировала способностью удерживать в фокусе большое поле зрения. Другим подарком природы стал гребешок (цифра 6), обеспечивающий острое зрение. Глазной гребень отсутствует у млекопитающих и слабо развит у рептилий. Первое описание гребня датируется 1676 годом, и до сих пор этот орган остается предметом многочисленных исследований; и в то же время он слабо изучен. Его основная функция остается невыясненной; доподлинно известно только, что его формы и размеры у разных птиц различна. Основная функция гребня — снабжение стекловидного тела и внутренних слоев сетчатки кислородом и удаление продуктов метаболизма.
Основная функция гребня — снабжение стекловидного тела и внутренних слоев сетчатки кислородом и удаление продуктов метаболизма.
«Энциклопедический словарь Брокгауза и Ефрона»
Гребень (или гребешок) — малоизученный орган складчатой ткани, который находится под сетчаткой. Он хорошо снабжен кровеносными сосудами и, вероятно, питает сетчатку, а также защищает ее от ослепительного света или помогает обнаруживать движущиеся объекты. Глазной гребень обильно наполнен гранулами меланина, которые поглощают рассеянный свет, попадающий в глаза птицы, уменьшая фоновые блики. Небольшое потепление гребня вследствие поглощения света гранулами меланина предположительно повышает интенсивность метаболизма гребня, увеличивая секрецию питательных веществ в стекловидное тело. Вероятно, очень высокая ферментативная активность щелочной фосфатазы в глазном гребне поддерживает его высокую секреторную функцию для дополнительного питания сетчатки. Складки гребня сетчатки глаза птиц представляют собой богатую сосудистую сеть. Интенсивный транспортный обмен метаболитами между кровью, внутриглазной жидкостью и структурами гребня сетчатки происходит путем активной диффузии [14–16].
Интенсивный транспортный обмен метаболитами между кровью, внутриглазной жидкостью и структурами гребня сетчатки происходит путем активной диффузии [14–16].
Анализы ДНК показали, что человек… не венец эволюции
Анализ ДНК современных видов животных и их предшественников показал, что на ранних этапах эволюции позвоночные имели четыре типа колбочек. Однако в процессе эволюции птицы и рептилии сохранили себе тетрахроматичное зрение, тогда как общий предок млекопитающих во время царствования динозавров вел преимущественно ночной образ жизни и лишился чувствительности к ультрафиолету и синему свету. Далее, человечество должно сказать спасибо метеориту, погубившему динозавров! Ведь именно благодаря ему вымерли динозавры, и млекопитающие стремительно увеличили свое многообразие. И далее, в процессе дупликации и последующей мутации гена одного из оставшихся фоточувствительных пигментов некоторые виды млекопитающих (в том числе приматы) приобрели трихроматичное зрение (рис. 6).
Рисунок 6. Тимоти Голдсмитт в своей статье рассказывает, что в процессе эволюции птицы сохранили зрительные пигменты нашего общего с ними предка. Однако в процессе эволюции млекопитающие суши сначала утратили пигмент ультрафиолетового цвета, затем утратили пигмент синего цвета, став дихроматами. Далее в процессе эволюции разные виды благодаря мутациям восстанавливали трихроматичное зрение (в частности, приматы). Каждый тип колбочек представлен в виде треугольника.
«В мире науки»
Палочки, колбочки, а вы кто?
Сетчатка птиц имеет три типа фоторецепторов:
- Палочки обеспечивают черно-белое зрение при низкой освещенности.
- Колбочки — цветное зрение при ярком свете.
- Двойные колбочки — цветное зрение.
Птицы являются тетрахроматами и воспринимают ультрафиолет (рис. 7). Помимо этого, сетчатка птиц не имеет собственных сосудов, и это уменьшает аберрации высшего порядка, которые снижают качество зрения человека. Острое зрение птиц достигается за счет большого количества фоторецепторов. Для сравнения, максимально достижимая острота зрения зависит от плотности фоторецепторов на сетчатке глаза: у человека их количество составляет около 200 000 на 1 мм2, а у домашнего воробья — 400 000 на 1 мм2! Помимо этого, хищные птицы и птицы, способные развивать высокую скорость полета (ласточки) или «летать на месте» (колибри, зимородок), имеют вторую ямку для улучшенного бокового обзора. Другое главное преимущество птиц заключается в быстрой обработке информации во время полета. Чтобы «думать в полете», скорость восприятия зрения составляет от 130 до 150 Гц, в то время, как у человека — только 50 Гц [17]. С другой стороны, для «одомашненных» птиц подобная скорость восприятия света будет скорее помехой, чем преимуществом, ведь частота мерцания люминесцентной лампы составляет 50–60 Гц, поэтому для человека ее свет кажется непрерывным. А теперь представьте, как чувствует себя птица!
Острое зрение птиц достигается за счет большого количества фоторецепторов. Для сравнения, максимально достижимая острота зрения зависит от плотности фоторецепторов на сетчатке глаза: у человека их количество составляет около 200 000 на 1 мм2, а у домашнего воробья — 400 000 на 1 мм2! Помимо этого, хищные птицы и птицы, способные развивать высокую скорость полета (ласточки) или «летать на месте» (колибри, зимородок), имеют вторую ямку для улучшенного бокового обзора. Другое главное преимущество птиц заключается в быстрой обработке информации во время полета. Чтобы «думать в полете», скорость восприятия зрения составляет от 130 до 150 Гц, в то время, как у человека — только 50 Гц [17]. С другой стороны, для «одомашненных» птиц подобная скорость восприятия света будет скорее помехой, чем преимуществом, ведь частота мерцания люминесцентной лампы составляет 50–60 Гц, поэтому для человека ее свет кажется непрерывным. А теперь представьте, как чувствует себя птица!
Рисунок 7. Цветовое восприятие птичьего зрения. Схематически пространство цветового зрения человека можно изобразить в виде области плоского треугольника (слева). На треугольнике отложены длины волн, воспринимаемых каждым из типов колбочек. Чтобы отразить цветовое зрение птицы, необходимо добавить еще одну ось к треугольнику (справа), которая будет отражать наложение от ультрафиолета на видимый спектральный диапазон. В результате получится тетраэдр: цвета в присутствии УФ отмечены плавающими шарами над основанием тетраэдра. Чем выше доля УФ в спектре, тем выше расположены точки. Например, оперение расписного овсянкового кардинала (внизу слева) отражает различное количество ультрафиолета в дополнение к цветам, различимых для человеческого глаза. Внизу справа изображен спектр отражения его оперения в видимом и УФ-спектре. Именно в УФ-области будут заметны различия по рисунку оперения.
Цветовое восприятие птичьего зрения. Схематически пространство цветового зрения человека можно изобразить в виде области плоского треугольника (слева). На треугольнике отложены длины волн, воспринимаемых каждым из типов колбочек. Чтобы отразить цветовое зрение птицы, необходимо добавить еще одну ось к треугольнику (справа), которая будет отражать наложение от ультрафиолета на видимый спектральный диапазон. В результате получится тетраэдр: цвета в присутствии УФ отмечены плавающими шарами над основанием тетраэдра. Чем выше доля УФ в спектре, тем выше расположены точки. Например, оперение расписного овсянкового кардинала (внизу слева) отражает различное количество ультрафиолета в дополнение к цветам, различимых для человеческого глаза. Внизу справа изображен спектр отражения его оперения в видимом и УФ-спектре. Именно в УФ-области будут заметны различия по рисунку оперения.
«В мире науки»
Так же, как и у млекопитающих, у дневных видов птиц палочек всего один вид, но целых шесть видов колбочек (для сравнения: у человека и приматов — три вида, а других млекопитающих — два). Четыре вида колбочек, максимально чувствительных к фиолетовой (ультрафиолетовой), синей, зеленой и красной областям спектра, обеспечивают птицам богатое цветное зрение (рис. 7). Оставшиеся два вида соединены вместе и функционируют как единый фоторецептор. Они называются двойными колбочками, и их роль состоит в восприятии не цвета, а движущихся объектов. Количество разных колбочек не одинаково. В сетчатке бóльшую долю среди фоторецепторов занимают двойные колбочки (40,7%), затем зеленые (21,1%), красные (17,1%), синие (12,6%) и фиолетовые (8,5%) (рис. 8).
Четыре вида колбочек, максимально чувствительных к фиолетовой (ультрафиолетовой), синей, зеленой и красной областям спектра, обеспечивают птицам богатое цветное зрение (рис. 7). Оставшиеся два вида соединены вместе и функционируют как единый фоторецептор. Они называются двойными колбочками, и их роль состоит в восприятии не цвета, а движущихся объектов. Количество разных колбочек не одинаково. В сетчатке бóльшую долю среди фоторецепторов занимают двойные колбочки (40,7%), затем зеленые (21,1%), красные (17,1%), синие (12,6%) и фиолетовые (8,5%) (рис. 8).
У птиц двойные колбочки представляют собой две плотно сросшиеся колбочки неравного размера. Их функция, по сравнению с обычными колбочками, до сих пор неясна, но главным образом они задействованы в ахроматических задачах — то есть не связаны с восприятием цветового зрения, а отвечают за регистрацию движения, определение яркости и поляризацию [10].
Один за всех и все за одного!
Родопсин и ретиналь — единственные молекулы, которые отвечают за зрение у всех позвоночных. Тогда возникает вопрос: почему же все животные видят по-разному? И почему палочки и колбочки различны? Неужели нельзя было сделать так, чтобы одна колбочка воспринимала сразу все цвета, зачем такое большое разнообразие? Дело в том, что все известные эволюционно различные классы зрительных пигментов, хоть и связаны с универсальным хромофором — 11-цис-ретиналем, — но имеют поглощение на разных длинах волн. Разница в их восприятии связана с различиями в белковой части пигмента — опсине (у птиц — один для палочек и четыре для колбочек) (таблица 1). Рекордсменом животного мира по количеству фоторецепторов по праву считается рак-богомол — он имеет их 16 штук! Следом за ним расположилась бабочка Sarpedon nipponum с 15 типами фоторецепторов. При этом известно, что количество фоторецепторов не оказывает существенного влияния на восприимчивость света за пределами тетрахроматии, а помогает лишь быстрее определять цвета. Подробнее с этими чудесами можно ознакомиться в статье «Биомолекулы» «Разные глаза — разные возможности» [18].
Тогда возникает вопрос: почему же все животные видят по-разному? И почему палочки и колбочки различны? Неужели нельзя было сделать так, чтобы одна колбочка воспринимала сразу все цвета, зачем такое большое разнообразие? Дело в том, что все известные эволюционно различные классы зрительных пигментов, хоть и связаны с универсальным хромофором — 11-цис-ретиналем, — но имеют поглощение на разных длинах волн. Разница в их восприятии связана с различиями в белковой части пигмента — опсине (у птиц — один для палочек и четыре для колбочек) (таблица 1). Рекордсменом животного мира по количеству фоторецепторов по праву считается рак-богомол — он имеет их 16 штук! Следом за ним расположилась бабочка Sarpedon nipponum с 15 типами фоторецепторов. При этом известно, что количество фоторецепторов не оказывает существенного влияния на восприимчивость света за пределами тетрахроматии, а помогает лишь быстрее определять цвета. Подробнее с этими чудесами можно ознакомиться в статье «Биомолекулы» «Разные глаза — разные возможности» [18].
| Пигмент | Значение | Длина волны (λmax, нм) | Наличие у человека, % | Наличие у птиц, % |
|---|---|---|---|---|
| Rh2 (Rhodopsin) | Родопсин палочек. Отвечает за ночное зрение, не чувствителен к цвету | 498 | 100 | 100 |
| LWS (Long wavelength sensitive) | Родопсин «красных» колбочек, длинноволновая область | 500–570 | 64 | 17,1 |
| MWS/Rh3 (Medium wavelength sensitive). (У млекопитающих отсутствует Rh3 родопсин, но вместо него есть MWS) | Родопсин «зеленых» колбочек, средние длины волн. | 480–530 | 32 | 21,1 |
| SWS1 (Short wavelength sensitive type 1) | Родопсин «синих» колбочек, малые длины волн | 400–470 | 4 | 12,6 |
| SWS2 (Short wavelength sensitive type 2) | Родопсин «ультрафиолетовых» колбочек, коротковолновая область | 354–445 | — | 8,5 |
| LWS + пигмент (A) | Двойная колбочка, воспринимает не цвета, а движущиеся объекты | Точные данные по восприятию цвета отсутствуют | — | 40,7 |
Рисунок 8А. Зрительные пигменты колбочек сетчатки человека. Цветное зрение приматов трихроматично (то есть определяется тремя типами колбочек, каждая из которых содержит свой тип светочувствительного пигмента). Справа указана доля колбочек в глазу.
Справа указана доля колбочек в глазу.
Рисунок 8Б. Зрительные пигменты колбочек сетчатки у птиц. Глаз птиц четыреххроматичен и способен различить большее количество цветов.
«Википедия»
Рисунок 8В. Оперение и части тела птиц по-разному отражают УФ-свет. Причина, по которой птицам выгодно видеть в УФ-диапазоне, весьма банальна — так проще отыскивать корм. Плоды и ягоды отражают УФ-лучи, что делает их более заметными для птиц (поэтому дрозды так любят лакомиться труднодоступной ягодой), а хищные птицы видят тропинки мышей — они помечены отражающими УФ мочой и экскрементами. Помимо этого, по интенсивности отражения УФ от оперения можно определить, молодое перо или старое. Так, у совы в УФ-освещении только крайние перья крыла имеют яркую флуоресценцию — именно они молодые, а все остальные будут заменены в ближайшее время. Однако обладатели самого лучшего цветовосприятия среди наземных позвоночных лишаются своего исключительного зрения в сумерках — дневным птицам требуется в 5–20 раз больше света, чем людям для ориентации в темноте.
Однако обладатели самого лучшего цветовосприятия среди наземных позвоночных лишаются своего исключительного зрения в сумерках — дневным птицам требуется в 5–20 раз больше света, чем людям для ориентации в темноте.
«Химия и жизнь», Bird fellow
Рисунок 8Г. У тупиков, пингвинов и некоторых других видов птиц в УФ-диапазоне отражает клюв, что, вероятно также связано с необходимостью распознавать партнера среди множества одинаковых птиц [20].
Рисунок 8Д. Некоторые птицы специально меняют оперение в период размножения на более УФ-отражающее, чтобы казаться более привлекательными. Также птицы имеют уникальный рисунок в УФ-диапазоне, что позволяет им лучше распознавать партнера.
«Химия и жизнь»
Встроенный светофильтр
Колбочки птиц, рыб и других позвоночных сохранили некоторые особенности, утерянные млекопитающими. Из них самое важное для цветового зрения — это наличие цветных капель жира. В зависимости от их типа, колбочки содержат красные, желтые, почти бесцветные и прозрачные липидные капельки. Цвет липидной капли определяется содержанием липидов, каротиноидов и специальным компонентом, образуя вместе природный светофильтр разного размера (рис. 9). При прохождении через липидную каплю отсекается часть длин волн, на которые реагируют колбочки, благодаря чему птицы способны различать большее количество цветов. Предполагается, что липидная капля образуется в результате жирового перерождения митохондрии во время дискогенеза или немного позже, после чего происходит их слияние и образование капли перед базальными дисками [10].
Из них самое важное для цветового зрения — это наличие цветных капель жира. В зависимости от их типа, колбочки содержат красные, желтые, почти бесцветные и прозрачные липидные капельки. Цвет липидной капли определяется содержанием липидов, каротиноидов и специальным компонентом, образуя вместе природный светофильтр разного размера (рис. 9). При прохождении через липидную каплю отсекается часть длин волн, на которые реагируют колбочки, благодаря чему птицы способны различать большее количество цветов. Предполагается, что липидная капля образуется в результате жирового перерождения митохондрии во время дискогенеза или немного позже, после чего происходит их слияние и образование капли перед базальными дисками [10].
Рисунок 9А. Расположение колбочек с липидными каплями в сетчатке цыпленка. На микрофотографии сетчатки гаички хорошо заметны бесцветные, желтые и красные пятна. Все окрашенные капли служат светофильтрами, не пропускающими короткие длины волн. Зрительные пигменты SWS1 (воспринимают ультрафиолет) содержат самые маленькие, бесцветные капельки T (transparent) и обладают максимумом поглощения при 400 нм. Эти капли обладают голубовато-белой флуоресценцией в ультрафиолетовом свете (второе изображение слева). Размер самых маленьких капель составляет 1–2 мкм, а самых больших — около 5 мкм. Доподлинно известно, что цвет липидной капли может меняться с возрастом птицы, однако причины этого явления еще неясны [10], [21].
Эти капли обладают голубовато-белой флуоресценцией в ультрафиолетовом свете (второе изображение слева). Размер самых маленьких капель составляет 1–2 мкм, а самых больших — около 5 мкм. Доподлинно известно, что цвет липидной капли может меняться с возрастом птицы, однако причины этого явления еще неясны [10], [21].
Рисунок 9Б. Благодаря липидным каплям птицы видят и различают гораздо большее количество оттенков цветов — по сравнению с человеком. В шуточной форме это можно изобразить в виде незамысловатой картинки.
коллаж автора
Организованный хаос
Большинство опытов по изучению глаз птиц проводится на цыплятах или взрослых курицах. Колбочки расположены на сетчатке неравномерно, неупорядоченно, но и не хаотично. Относительно друг друга они выстраиваются в сложную мозаику, причем вокруг колбочки одного цвета могут находиться колбочки только другого (но не того же самого) цвета. То есть, при наличии в глазу шести видов колбочек, сетчатку можно разбить на шесть видов сегментов мозаик, чтобы увидеть упорядоченность (рис. 10). Закономерность, найденная в сетчатке цыплят, оказалась верной и для других птиц [22].
То есть, при наличии в глазу шести видов колбочек, сетчатку можно разбить на шесть видов сегментов мозаик, чтобы увидеть упорядоченность (рис. 10). Закономерность, найденная в сетчатке цыплят, оказалась верной и для других птиц [22].
Рисунок 10. Организованный хаос. Цветные точки в квадрате слева соответствуют расположению зеленых, синих, красных, фиолетовых и двойных (обозначено черным) фоторецепторов сетчатки курицы. На первый взгляд кажется, что колбочки перемешаны между собой беспорядочно, но на самом деле они имеют упорядоченность относительно колбочек своего вида, независимо от колбочек другого цвета. Таким образом, колбочка формирует вокруг себя «зону исключения» для колбочки своего же вида (обозначено закрашенными кружками на рисунках справа). При этом в «зоне исключения» могут располагаться колбочки всех других типов. Это распределение подчинено пространственному разделению. Каждый набор колбочек нельзя назвать идеально однородным, но он однороден настолько, что может иметь ограничения по распределению пяти различных колбочек. Гиперупорядоченость, в свою очередь, образует гипероднородность — то есть, если разбить сетчатку на сегменты условного размера, то количество всех рецепторов внутри каждого сегмента не будет сильно различаться.
Гиперупорядоченость, в свою очередь, образует гипероднородность — то есть, если разбить сетчатку на сегменты условного размера, то количество всех рецепторов внутри каждого сегмента не будет сильно различаться.
Quanta magazine
Мигательная перепонка
Еще одно преимущество птиц, которое они сохранили (в отличие от млекопитающих) в процессе эволюции, — мигательная перепонка (рис. 11). Она представляет собой (полу)прозрачное третье веко, существующее у некоторых животных и позволяющее защищать глаз или увлажнять его без потери видимости. У человека она осталась в виде рудимента — третьего века в уголке глаза.
Рисунок 11. Мигательная перепонка (третье веко) у птиц и человека.
«Википедия»
Почему у Тук-Тукыча глаза целые?
Дятлу можно позавидовать — работая целыми днями напролет в поисках пищи, он точно никогда не испытывал головной боли или мигрени. Как же это возможно? Впервые полностью расшифровать механизм предохранения головы дятла от сотрясений удалось двум американским ученым — Айвану Швобу из Калифорнийского университета в Дэвисе и Филиппу Мэю из Калифорнийского университета в Лос-Анджелесе. За эту работу они получили Шнобелевскую премию 2006 года по орнитологии! Помимо сверхтвердого клюва, полной блокировки мышц шеи и полностью перпендикулярных поверхности ударов, было показано, что дятел имеет систему безопасности для сохранности глаз. Во время ударов мигательная перепонка опускается на глаза птицы, чтобы уберечь глазное яблоко от вибраций и не допустить отслоения сетчатки [23].
За эту работу они получили Шнобелевскую премию 2006 года по орнитологии! Помимо сверхтвердого клюва, полной блокировки мышц шеи и полностью перпендикулярных поверхности ударов, было показано, что дятел имеет систему безопасности для сохранности глаз. Во время ударов мигательная перепонка опускается на глаза птицы, чтобы уберечь глазное яблоко от вибраций и не допустить отслоения сетчатки [23].
Вы не подскажете, как лететь на юг?
Некоторые виды птиц, в зависимости от времени года, совершают миграции к местам зимовок и обратно. Как им удается точно распознавать, где север, без карманного компаса? Помимо способности определять положение Солнца и звезд [24], природа снабдила перелетных птиц уникальным приспособлением, расположенным прямо в их глазу. Детали каждого из устройств не до конца ясны, но больше всего внимания ученые уделяют магнитному компасу, который по праву занимает главенствующую позицию среди имеющихся у птиц средств навигации. Следом за ним следует солнечный компас, принцип действия которого основан на чувствительности к поляризованному свету [25].
Некоторые виды птиц способны использовать для вычисления направления магнитного поля Земли квантовый компьютер, встроенный прямо в их глаза. Но прежде всего, стоит понимать, что птичий компас отличен от человеческого устройства — если привычный нам прибор указывает стрелкой на северный и южный магнитные полюса, то птичий навигатор будет определять положение «магнитного полюса» и «магнитного экватора». На самом деле, птицы реагируют на магнитное наклонение — угол, образуемый силовыми линиями поля с плоскостью горизонта, и им совершенно безразлично, где север, а где юг. Другая важная составляющая магнитного компаса птиц — освещение. Птичий навигатор полностью отказывается работать в темноте: ему требуется, пусть и слабое, освещение с длиной волны 400–570 нм. Более подробно о системе навигации пернатых можно прочитать в статье «Биомолекулы» «Пазл магнитного компаса птиц» [26].
Большинство исследователей полагает, что перелетные птицы способны воспринимать магнитное поле Земли благодаря процессам магниторецепции криптохрома.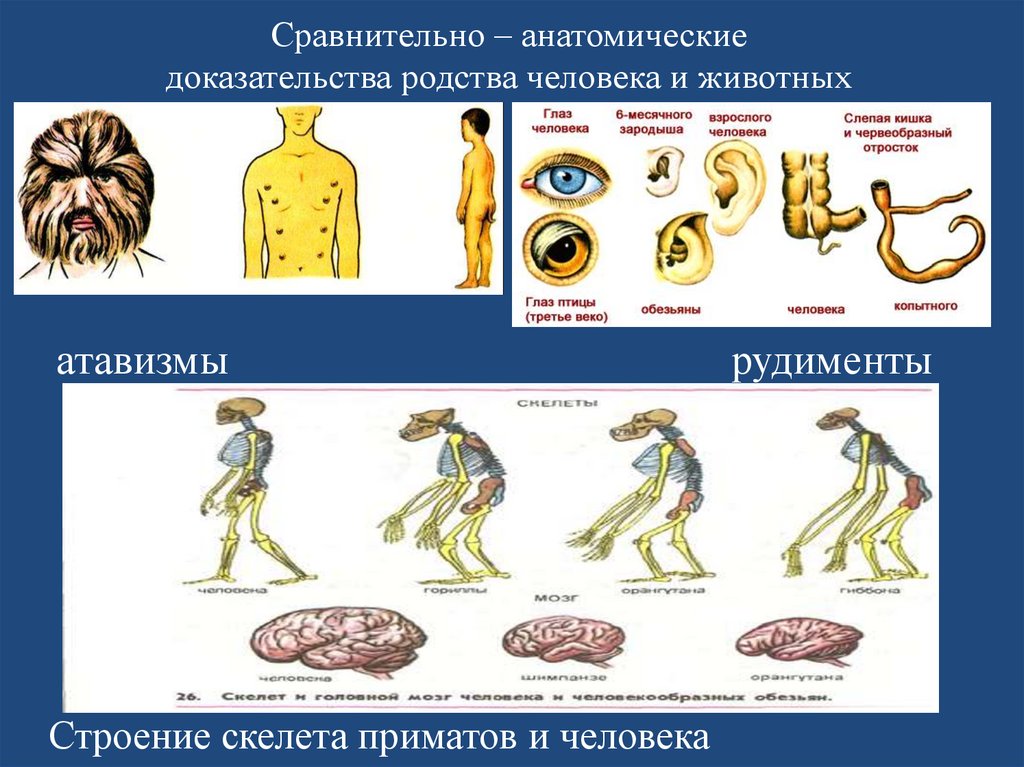 Птицы поворачивают голову, чтобы определить магнитное поле. В глазу перелетной птицы расположены чувствительные к свету белки криптохромы со светособирающим хромоформом птерином. Криптохром-4 выглядит самым подходящим претендентом на роль руководителя компаса. Во-первых, он экспрессируется сезонно (весной и осенью), а во-вторых, он расположен не везде, а только в красных и двойных колбочках.
Птицы поворачивают голову, чтобы определить магнитное поле. В глазу перелетной птицы расположены чувствительные к свету белки криптохромы со светособирающим хромоформом птерином. Криптохром-4 выглядит самым подходящим претендентом на роль руководителя компаса. Во-первых, он экспрессируется сезонно (весной и осенью), а во-вторых, он расположен не везде, а только в красных и двойных колбочках.
Три консервативных аминокислотных остатка триптофана (Trp 400, Trp 377 и Trp 324; рис. 12) криптохрома являются ключевыми элементами: под действием синего света происходит перераспределение заряда между этой триадой и флавином. В результате образуется устойчивая радикальная пара, чувствительная к магнитному полю. Но даже это простое объяснение вызывает ряд вопросов, ведь криптохром поглощает в диапазоне 400–500 нм [27], в то время как птицы продолжают безошибочно ориентироваться и при более длинноволновом освещении.
Более подробно о магниторецепции и работе криптохрома можно прочитать в статье «Биомолекулы» «Нобель vs Шнобель, или Механизмы магниторецепции» [28].
Немного о квантах света
Принцип запрета Паули гласит, что два электрона на одной орбите представляют собой волны, равновероятно направленные (в смысле их спинов) в противоположных направлениях. Квантовая запутанность — это особое явление, при котором квантовые состояния объектов взаимозависимы — две частицы устанавливают связь друг с другом и утрачивают индивидуальность. Например, при изменении спина одной из двух созависимых частиц обязательно изменится спин и второй частицы [29].
Рисунок 12. Магниторецепция птиц построена на принципе Паули и связана с фотопревращениями молекул криптохрома, связанных с оптическим нервом птицы. Под воздействием синего света криптохром перераспределяет свой заряд между триадой триптофанов и флавином, становясь чувствительным к магнитному полю. Далее, при попадании на образовавшийся радикал электрона с «правильным» спином, птица чувствует магнитное поле. Таким образом, образовавшийся радикал становится ловушкой для электронов.
«Нейроновости»
Чувствительность птиц к изменениям магнитного поля весьма высока. Так, в экспериментах с европейскими малиновками изменение магнитного поля на 15 нТл (менее одной тысячной силы магнитного поля Земли) оказалось достаточным, чтобы нарушить чувство направления птицы [30]. Другое исследование показало, что состояние квантовой запутанности в глазах птицы должно длиться как минимум 100 мкс, чтобы птица чувствовала направление магнитного поля. На настоящее время ученым удалось создать установку, поддерживающую квантовую запутанность только 80 мкс, а птицы могут поддерживать это состояние благодаря затухающему осцилляторному колебанию электронов при их встрече с клетками глаза [31]!
Согласно гипотезе магниторецепции, молекулы криптохрома способны провзаимодействовать только с электронами с противоположным спином. Таким образом, если спины останутся в запутанном состоянии, это приведет к получению глазом энергии от фотона. Но если птица ориентирована «неправильно», то спины не становятся параллельными, не рекомбинируют, и оптический нерв не получит возбуждающего стимула. Согласно другому представлению, свет возбуждает криптохромы, за счет чего происходит испускание непарных электронов, взаимодействующих с магнитным полем Земли. То есть, существуют магниточувствительные бирадикальные химические реакции, которые чувствительны к расположению птицы в магнитном поле Земли, или, иными словами — чувствительны к углу между силовыми линиями магнитного поля и поверхностью Земли [32–35].
Но если птица ориентирована «неправильно», то спины не становятся параллельными, не рекомбинируют, и оптический нерв не получит возбуждающего стимула. Согласно другому представлению, свет возбуждает криптохромы, за счет чего происходит испускание непарных электронов, взаимодействующих с магнитным полем Земли. То есть, существуют магниточувствительные бирадикальные химические реакции, которые чувствительны к расположению птицы в магнитном поле Земли, или, иными словами — чувствительны к углу между силовыми линиями магнитного поля и поверхностью Земли [32–35].
В настоящее время о зрении птиц известно очень мало, но в то же время, уже известные факты ставят ученых в тупик, а решение поставленных задач может служить толчком к развитию новых технологий!
- Зрительный родопсин — рецептор, реагирующий на свет;
- Вавилов С. И. Глаз и солнце. М.: «Академия наук СССР», 1927. — 334 с.;
- Островский М. А., Фельдман Т. Б. Фотобиология и фотохимия первичных процессов зрения.
 Дубна : Междунар. ун-т природы, о-ва и человека «Дубна», 2011. — 58 с.;
Дубна : Междунар. ун-т природы, о-ва и человека «Дубна», 2011. — 58 с.; - Как исследовать клетку на уровне отдельных биомолекул;
- 12 методов в картинках: микроскопия;
- Крупные подробности микроскопического мира: Нобелевская премия по химии 2017;
- Островский М. А. (2005). Молекулярные механизмы повреждающего действия света на структуры глаза и системы защиты от такого повреждения. Успехи биологической химии. 45, 173–204;
- Зрительный родопсин — рецептор, реагирующий на свет;
- Аллостерические регуляторы GPCR: ключи от всех замков;
- Хохлова Т.В. (2009). Фоторецепторы птиц: молекулярная генетика зрительных пигментов, структурные и функциональные особенности клеток и их топография. Сенсорные системы. 23, 91–105;
- Разные глаза — разные возможности;
- Откуда пошло зрение;
- Terry M.
 Tritt, M. A. Subramanian. (2006). Thermoelectric Materials, Phenomena, and Applications: A Bird’s Eye View. MRS Bull.. 31, 188-198;
Tritt, M. A. Subramanian. (2006). Thermoelectric Materials, Phenomena, and Applications: A Bird’s Eye View. MRS Bull.. 31, 188-198; - Шляпникова А. А. (2012). Морфологические особенности сетчатки глаза у птиц (куры, утки, гуси). Ученые записки Казанской государственной академии ветеринарной медицины им. Н.Э Баумана. 1, 212–216;
- Дегтярев В. В., Шляпникова А. А. (2015). Результаты электронно-микроскопических исследований ультраструктуры гребня сетчатки глаза птиц. Современные проблемы науки и образования. 3, 604–605;
- Соломахина Л. А. (2016). Нормальные вариации глазного дна птиц. VetPharma. 3, 56–65;
- Jannika E. Boström, Marina Dimitrova, Cindy Canton, Olle Håstad, Anna Qvarnström, Anders Ödeen. (2016). Ultra-Rapid Vision in Birds. PLoS ONE. 11, e0151099;
- Разные глаза — разные возможности;
- Lukas Hofmann, Krzysztof Palczewski.
 (2015). Advances in understanding the molecular basis of the first steps in color vision. Progress in Retinal and Eye Research. 49, 46-66;
(2015). Advances in understanding the molecular basis of the first steps in color vision. Progress in Retinal and Eye Research. 49, 46-66; - Evtukh G. (2019). Fluorescence among Fraterculinae subfamily. Русский орнитологический журнал. 28, 2134–2142;
- N. S. Hart, J. C. Partridge, I. C. Cuthill, A. T. D. Bennett. (2000). Visual pigments, oil droplets, ocular media and cone photoreceptor distribution in two species of passerine bird: the blue tit ( Parus caeruleus L.) and the blackbird ( Turdus merula L.). Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology. 186, 375-387;
- Yang Jiao, Timothy Lau, Haralampos Hatzikirou, Michael Meyer-Hermann, Joseph C. Corbo, Salvatore Torquato. (2014). Avian photoreceptor patterns represent a disordered hyperuniform solution to a multiscale packing problem. Phys. Rev. E. 89;
- Schwab I.
 R. (2002). Cure for a headache. Br J Ophthalmol. 86, 843;
R. (2002). Cure for a headache. Br J Ophthalmol. 86, 843; - Н. С. Чернецов, N. S. Chernetsov. (2016). Ориентация и навигация мигрирующих птиц. Зоол. ж.. 95, 128-146;
- Зуева Л. В., Макаров В. И., Голубева Т. Б., Инюшин М. Ю. (2015). В поисках структурного базиса поляризационной чувствительности у птиц. Энергетика и годовые циклы птиц. 1, 126–130;
- Пазл магнитного компаса птиц;
- Л. А. Астахова, А. Ю. Ротов, К. В. Кавокин, Н. С. Чернецов, М. Л. Фирсов. (2019). Cвязь магнитного компаса и фоторецепции у птиц: гипотезы и нерешенные вопросы. Журн. общ. биологии. 80, 83-94;
- Нобель vs Шнобель, или Механизмы магниторецепции;
- PhilipR.A. May, Paul Newman, JoaquinM. Fuster, Ada Hirschman. (1976). WOODPECKERS AND HEAD INJURY. The Lancet. 307, 454-455;
- Thorsten Ritz, Roswitha Wiltschko, P.
 J. Hore, Christopher T. Rodgers, Katrin Stapput, et. al.. (2009). Magnetic Compass of Birds Is Based on a Molecule with Optimal Directional Sensitivity. Biophysical Journal. 96, 3451-3457;
J. Hore, Christopher T. Rodgers, Katrin Stapput, et. al.. (2009). Magnetic Compass of Birds Is Based on a Molecule with Optimal Directional Sensitivity. Biophysical Journal. 96, 3451-3457; - Erik M. Gauger, Elisabeth Rieper, John J. L. Morton, Simon C. Benjamin, Vlatko Vedral. (2011). Sustained Quantum Coherence and Entanglement in the Avian Compass. Phys. Rev. Lett.. 106;
- Кишкинёв Д. А.,Чернецов Н. С. (2014). Магниторецепторные системы у птиц: обзор современных исследований. Журнал общей биологии. 75, 104–123;
- Jingjing Xu, Lauren E. Jarocha, Tilo Zollitsch, Marcin Konowalczyk, Kevin B. Henbest, et. al.. (2021). Magnetic sensitivity of cryptochrome 4 from a migratory songbird. Nature. 594, 535-540;
- Norman Ginsburg, Virginia Nilsson. (1971). MEASURING FLICKER THRESHOLDS IN THE BUDGERIGAR1. Journal of the Experimental Analysis of Behavior.
 15, 189-192;
15, 189-192; - Henrik Mouritsen, Gesa Feenders, Miriam Liedvogel, Wiebke Kropp. (2004). Migratory Birds Use Head Scans to Detect the Direction of the Earth’s Magnetic Field. Current Biology. 14, 1946-1949.
С точки зрения насекомого
: 22 Июл 2013 , «Но чтоб сие здание [Академия] непременно и полезно было, то имеет оное само себя править» , том 50, №2
Считается, что до 90% знаний о внешнем мире человек получает при помощи своего стереоскопического зрения. Зайцы обзавелись боковым зрением, благодаря которому они могут видеть объекты, находящиеся сбоку и даже позади себя. У глубоководных рыб глаза могут занимать до половины головы, а теменной «третий глаз» миноги позволяет ей неплохо ориентироваться в воде. Змеи способны видеть только движущийся объект, а самыми зоркими в мире признаны глаза сокола-сапсана, способного выследить добычу с высоты 8 км!
Змеи способны видеть только движущийся объект, а самыми зоркими в мире признаны глаза сокола-сапсана, способного выследить добычу с высоты 8 км!
Но как видят мир представители самого многочисленного и разнообразного класса живых существ на Земле – насекомых? Наряду с позвоночными животными, которым они проигрывают только по размерам тела, именно насекомые обладают наиболее совершенным зрением и сложноустроенными оптическими системами глаза. Хотя фасеточные глаза насекомых не обладают аккомодацией, вследствие чего их можно назвать близорукими, однако они, в отличие от человека, способны различать чрезвычайно быстро двигающиеся объекты. А благодаря упорядоченной структуре своих фоторецепторов многие из них обладают настоящим «шестым чувством» – поляризационным зрением
Меркнет зрение – сила моя,
Два незримых алмазных копья…
А. Тарковский (1983)
Трудно переоценить значение света (электромагнитного излучения видимого спектра) для всех обитателей нашей планеты. Солнечный свет служит основным источником энергии для фотосинтезирующих растений и бактерий, а опосредованно через них – и для всех живых организмов земной биосферы. Свет непосредственно влияет на протекание всего многообразия жизненных процессов животных, от размножения до сезонной смены окраски. И, конечно, благодаря восприятию света специальными органами чувств, животные получают значительную (а часто и большую) часть сведений об окружающем мире, могут различать форму и цвет объектов, определять движение тел, ориентироваться в пространстве и т. п.
Солнечный свет служит основным источником энергии для фотосинтезирующих растений и бактерий, а опосредованно через них – и для всех живых организмов земной биосферы. Свет непосредственно влияет на протекание всего многообразия жизненных процессов животных, от размножения до сезонной смены окраски. И, конечно, благодаря восприятию света специальными органами чувств, животные получают значительную (а часто и большую) часть сведений об окружающем мире, могут различать форму и цвет объектов, определять движение тел, ориентироваться в пространстве и т. п.
Зрение особенно важно для животных, способных активно передвигаться в пространстве: именно с возникновением подвижных животных начал формироваться и совершенствоваться зрительный аппарат – сложнейший из всех известных сенсорных систем. К таким животным относятся позвоночные и среди беспозвоночных – головоногие моллюски и насекомые. Именно эти группы организмов могут похвалиться самыми сложноустроенными органами зрения.
Однако зрительный аппарат у этих групп значительно различается, как и восприятие образов. Считается, что насекомые в целом более примитивны по сравнению с позвоночными, не говоря уже о высшем их звене – млекопитающих, и, естественно, человеке. Но вот насколько различается их зрительное восприятие? Иными словами, намного ли отличается от нашего мир, увиденный глазами маленького создания по имени муха?
Мозаика из шестигранников
Зрительная система насекомых в принципе не отличается от таковой у других животных и состоит из периферических органов зрения, нервных структур и образований центральной нервной системы. Но что касается морфологии органов зрения, то здесь различия просто бросаются в глаза.
Всем знакомы сложные фасеточные глаза насекомых, которые встречаются у взрослых насекомых или у личинок насекомых, развивающихся с неполным превращением, т. е. без стадии куколки. Исключений из этого правила не так много: это блохи (отряд Siphonaptera), веерокрылые (отряд Strepsiptera), большинство чешуйниц (семейство Lepismatidae) и весь класс скрыточелюстных (Entognatha).
Фасеточный глаз по виду напоминает корзинку спелого подсолнуха: он состоит из набора фасеток (омматидиев) – автономных приемников светового излучения, имеющих все необходимое для регуляции светового потока и формирования изображения. Число фасеток сильно варьирует: от нескольких у щетинохвосток (отряд Thysanura) до 30 тыс. у стрекоз (отряд Aeshna). Удивительно, но число омматидиев может варьироваться даже внутри одной систематической группы: например, ряд видов жуков-жужелиц, обитающих на открытых пространствах, имеют хорошо развитые фасеточные глаза с большим количеством омматидиев, в то время как у жужелиц, обитающих под камнями, глаза сильно редуцированы и состоят из небольшого числа омматидиев.
Верхний слой омматидиев представлен роговицей (хрусталиком) – участком прозрачной кутикулы, секретируемой специальными клетками, которая представляет собой своеобразную шестигранную двояковыпуклую линзу. Под роговицей у большинства насекомых располагается прозрачный кристаллический конус, структура которого может различаться у разных видов. У некоторых видов, особенно ведущих ночной образ жизни, в светопреломляющем аппарате имеются дополнительные структуры, играющие главным образом роль антибликового покрытия и увеличивающие светопропускание глаза.
У некоторых видов, особенно ведущих ночной образ жизни, в светопреломляющем аппарате имеются дополнительные структуры, играющие главным образом роль антибликового покрытия и увеличивающие светопропускание глаза.
Изображение, сформированное хрусталиком и кристаллическим конусом, попадает на светочувствительные ретинальные (зрительные) клетки, представляющие собой нейрон с коротким хвостиком-аксоном. Несколько ретинальных клеток образуют единый цилиндрический пучок – ретинулу. Внутри каждой такой клетки на стороне, обращенной внутрь омматидия, расположен рабдомер – особое образование из множества (до 75—100 тыс.) микроскопических трубочек-ворсинок, в мембране которых содержится зрительный пигмент. Как и у всех позвоночных, этим пигментом является родопсин – сложный окрашенный белок. Благодаря огромной площади этих мембран фоторецепторный нейрон содержит большое количество молекул родопсина (например, у плодовой мушки Drosophila это число превышает 100 млн!).
Рабдомеры всех зрительных клеток, объединенные в рабдом, и являются светочувствительными, рецепторными элементами фасеточного глаза, а все ретинулы в совокупности составляют аналог нашей сетчатки.
Светопреломляющий и светочувствительный аппарат фасетки по периметру окружают клетки с пигментами, которые играют роль световой изоляции: благодаря им световой поток, преломляясь, попадает на нейроны только одного омматидия. Но так устроены фасетки в так называемых фотопических глазах, приспособленных к яркому дневному свету.
Для видов, ведущих сумеречный или ночной образ жизни, характерны глаза другого типа – скотопические. Такие глаза имеют ряд приспособлений к недостаточному световому потоку, например, очень большие рабдомеры. Кроме того, в омматидиях таких глаз светоизолирующие пигменты могут свободно мигрировать внутри клеток, благодаря чему световой поток может попадать на зрительные клетки соседних омматидиев. Этот феномен лежит в основе и так называемой темновой адаптации глаз насекомых – увеличении чувствительности глаза при недостаточном освещении.
Этот феномен лежит в основе и так называемой темновой адаптации глаз насекомых – увеличении чувствительности глаза при недостаточном освещении.
При поглощении рабдомерами фотонов света в ретинальных клетках генерируются нервные импульсы, которые по аксонам направляются в парные зрительные доли головного мозга насекомых. В каждой зрительной доле имеется по три ассоциативных центра, где и осуществляется переработка потока зрительной информации, одновременно идущей от множества фасеток.
От одного до тридцати
Согласно древним легендам, у людей некогда имелся «третий глаз», отвечающий за сверхчувственное восприятие. Доказательств этому нет, однако та же минога и другие животные, такие как ящерица-гаттерия и некоторые земноводные, имеют необычные светочувствительные органы в «неположенном» месте. И в этом смысле насекомые не отстают от позвоночных: помимо обычных фасеточных глаз у них встречаются небольшие дополнительные глазки – оцелли, расположенные на лобно-теменной поверхности, и стеммы – по бокам головы.
Оцелли имеются в основном у хорошо летающих насекомых: взрослых особей (у видов с полным превращением) и личинок (у видов с неполным превращением). Как правило, это три глазка, расположенные в виде треугольника, но иногда срединный либо два боковых могут отсутствовать. По строению оцелли сходны с омматидиями: под светопреломляющей линзой у них находится слой прозрачных клеток (аналог кристаллического конуса) и сетчатка-ретинула.
Стеммы можно обнаружить у личинок насекомых, развивающихся с полным превращением. Их число и расположение варьирует в зависимости от вида: с каждой стороны головы может располагаться от одного до тридцати глазков. У гусениц чаще встречается шесть глазков, расположенных так, что каждый из них имеет обособленное поле зрения.
В разных отрядах насекомых стеммы могут отличаться друг от друга по строению. Эти различия связаны, возможно, с их происхождением от разных морфологических структур. Так, число нейронов в одном глазке может составлять от нескольких единиц до нескольких тысяч. Естественно, это сказывается на восприятии насекомыми окружающего мира: если некоторые из них могут видеть лишь перемещение светлых и темных пятен, то другие способны распознавать размеры, форму и цвет предметов.
Так, число нейронов в одном глазке может составлять от нескольких единиц до нескольких тысяч. Естественно, это сказывается на восприятии насекомыми окружающего мира: если некоторые из них могут видеть лишь перемещение светлых и темных пятен, то другие способны распознавать размеры, форму и цвет предметов.
Как мы видим, и стеммы, и омматидии представляют собой аналоги одиночных фасеток, пусть и видоизмененные. Однако у насекомых имеются и другие «запасные» варианты. Так, некоторые личинки (особенно из отряда двукрылых) способны распознать свет даже при полностью затененных глазках с помощью фоточувствительных клеток, расположенных на поверхности тела. А некоторые виды бабочек имеют так называемые генитальные фоторецепторы.
Все такие фоторецепторные зоны устроены схожим образом и представляют собой скопление из нескольких нейронов под прозрачной (или полупрозрачной) кутикулой. За счет подобных дополнительных «глаз» личинки двукрылых избегают открытых пространств, а самки бабочек используют их при откладке яиц в затененных местах.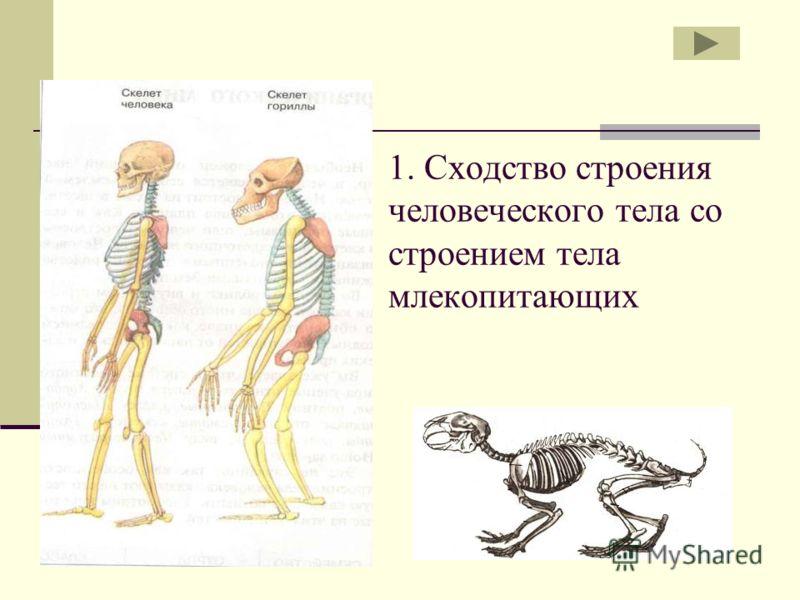
Фасеточный поляроид
На что способны сложноустроенные глаза насекомых? Как известно, у любого оптического излучения можно выделить три характеристики: яркость, спектр (длину волны) и поляризацию (ориентированность колебаний электромагнитной составляющей).
Спектральную характеристику света насекомые используют для регистрации и распознавания объектов окружающего мира. Практически все они способны воспринимать свет в диапазоне от 300—700 нм, в том числе и недоступную для позвоночных ультрафиолетовую часть спектра.
Как правило, разные цвета воспринимаются различными областями сложного глаза насекомых. Такая «локальная» чувствительность может различаться даже в пределах одного вида в зависимости от половой принадлежности особи. Нередко в одном и том же омматидии могут находиться различные цветовые рецепторы. Так, у бабочек рода Papilio два фоторецептора имеют зрительный пигмент с максимумом поглощения 360, 400 или 460 нм, еще два – 520 нм, а остальные – от 520 до 600 нм (Kelber et al. , 2001).
Но это далеко не все, что умеет глаз насекомого. Как упоминалось выше, в зрительных нейронах фоторецепторная мембрана микроворсинок рабдомера свернута в трубку круглого или гексагонального сечения. За счет этого часть молекул родопсина не участвуют в поглощении света из-за того, что дипольные моменты этих молекул располагаются параллельно ходу светового луча (Говардовский, Грибакин, 1975). В результате микроворсинка приобретает дихроизм – способность к различному поглощению света в зависимости от его поляризации. Повышению поляризационной чувствительности омматидия способствует и то, что молекулы зрительного пигмента не располагаются в мембране хаотично, как у человека, а ориентированы в одном направлении, да к тому же жестко закреплены.
Если глаз способен различить два источника света на основе их спектральных характеристик вне зависимости от интенсивности излучения, можно говорить о цветовом зрении. Но если он делает это, фиксируя поляризационный угол, как в данном случае, мы имеем все основания говорить о поляризационном зрении насекомых.
Как же воспринимают насекомые поляризованный свет? Исходя из структуры омматидия, можно предположить, что все фоторецепторы должны быть одновременно чувствительными как к определенной длине (длинам) световых волн, так и к степени поляризации света. Но в таком случае могут возникнуть серьезные проблемы – так называемое ложное восприятие цвета. Так, свет, отраженный с глянцевой поверхности листьев или водной глади, частично поляризуется. В этом случае мозг, анализируя данные фоторецепторов, может ошибиться в оценке интенсивности окраски либо формы отражающей поверхности.
Насекомые научились успешно справляться с подобными трудностями. Так, у ряда насекомых (в первую очередь мух и пчел) в омматидиях, воспринимающих только цвет, формируется рабдом закрытого типа, в котором рабдомеры не контактируют между собой. При этом у них имеются также омматидии с обычными прямыми рабдомами, чувствительные и к поляризационному свету. У пчел такие фасетки располагаются по краю глаза (Wehner, Bernard, 1993). У некоторых бабочек искажения при восприятии цвета снимаются за счет значительного искривления микроворсинок рабдомеров (Kelber et al., 2001).
У многих других насекомых, особенно у чешуекрылых, во всех омматидиях сохраняются обычные прямые рабдомы, поэтому их фоторецепторы способны одновременно воспринимать и «цветной», и поляризованный свет. При этом каждый из этих рецепторов чувствителен лишь к определенному поляризационному углу преференции и определенной длине световой волны. Такое сложное зрительное восприятие помогает бабочкам при питании и откладке яиц (Kelber et al., 2001).
Незнакомая Земля
Можно бесконечно углубляться в особенности морфологии и биохимии глаза насекомых и все равно затруднится в ответе на такой простой и одновременно невероятно сложный вопрос: как видят насекомые?
Человеку трудно даже представить образы, возникающие в головном мозге насекомых. Но все нужно заметить, что популярная сегодня мозаичная теория зрения, согласно которой насекомое видит изображение в виде своеобразного пазла из шестигранников, не совсем точно отражает суть проблемы. Дело в том, что хотя каждая единичная фасетка фиксирует отдельный образ, являющийся лишь частью цельной картины, эти изображения могут перекрываться с изображениями, полученными с соседних фасеток. Поэтому изображение мира, полученное с помощью огромного глаза стрекозы, состоящего из тысяч миниатюрных камер-фасеток, и «скромного» шестифасеточного глаза муравья, будет сильно различаться.
Что касается остроты зрения (разрешающей способности, т. е. способности различать степень расчлененности объектов), то у насекомых она определяется количеством фасеток, приходящихся на единицу выпуклой поверхности глаза, т. е. их угловой плотностью. В отличие от человека, глаза насекомых не обладают аккомодацией: радиус кривизны светопроводящей линзы у них не меняется. В этом смысле насекомых можно назвать близорукими: они видят тем больше деталей, чем ближе к объекту наблюдения находятся.
При этом насекомые с фасеточными глазами способны различать очень быстро движущиеся объекты, что объясняется высокой контрастностью и малой инерционностью их зрительной системы. К примеру, человек может различать лишь около двадцати вспышек в секунду, а пчела – в десять раз больше! Такое свойство жизненно важно для быстролетающих насекомых, которым нужно принимать решения непосредственно в полете.
Цветовые образы, воспринимаемые насекомыми, также могут быть гораздо сложнее и необычнее, чем у нас. К примеру, цветок, кажущийся нам белым, часто скрывает в своих лепестках множество пигментов, способных отражать ультрафиолетовый свет. И в глазах насекомых-опылителей он сверкает множеством красочных оттенков – указателей на пути к нектару.
Считается, что насекомые «не видят» красный цвет, который в «чистом виде» и встречается в прирорде чрезвычайно редко (исключение – тропические растения, опыляемые колибри). Однако цветы, окрашенные в красный цвет, часто содержат и другие пигменты, способные отражать коротковолновые излучения. А если учесть, что многие из насекомых способны воспринимать не три основных цвета, как человек, а больше (иногда до пяти!), то их зрительные образы должны представлять собой просто феерию красок.
И, наконец, «шестое чувство» насекомых – поляризационное зрение. С его помощью насекомым удается увидеть в окружающем мире то, о чем человек может получить лишь слабое представление с помощью специальных оптических фильтров. Насекомые же таким способом могут безошибочно определять местонахождение солнца на облачном небе и использовать поляризованный свет в качестве «небесного компаса». А водные насекомые в полете обнаруживают водоемы по частично поляризованному свету, отраженному от зеркала воды (Schwind, 1991). Но вот какие при этом они «видят» образы, человеку просто невозможно себе представить…
У всех, кто по той или иной причине интересуется зрением насекомых, может возникнуть вопрос: почему у них не сформировался камерный глаз, подобный человеческому глазу, со зрачком, хрусталиком и прочими приспособлениями?
На этот вопрос в свое время исчерпывающе ответил выдающийся американский физик-теоретик, Нобелевский лауреат Р. Фейнман: «Этому мешает несколько довольно интересных причин. Прежде всего, пчела слишком мала: если бы она имела глаз, похожий на наш, но соответственно уменьшенный, то размер зрачка оказался бы порядка 30 мкм, а поэтому дифракция была бы столь велика, что пчела все равно не могла бы видеть лучше. Слишком маленький глаз — это не очень хорошо. Если же такой глаз сделать достаточного размера, то он должен быть не меньше головы самой пчелы. Ценность сложного глаза в том и состоит, что он практически не занимает места – просто тоненький слой на поверхности головы. Так что, прежде чем давать советы пчеле, не забывайте, что у нее есть свои собственные проблемы!»
Поэтому неудивительно, что насекомые выбрали свой путь в зрительном познании мира. Да и нам, чтобы видеть его с точки зрения насекомых, пришлось бы, для сохранения привычной остроты зрения, обзавестись громадными фасеточными глазами. Вряд ли такое приобретение оказалось бы нам полезным с точки зрения эволюции. Каждому – свое!
Литература
Тыщенко В. П. Физиологияя насекомых. М.: Высшая школа, 1986, 304 С.
Klowden M. J. Physiological Systems in Insects. Academ Press, 2007. 688 p.
Nation J. L. Insect Physiology and Biochemistry. Second Edition: CRC Press, 2008.
В публикации использованы фото автора
: 22 Июл 2013 , «Но чтоб сие здание [Академия] непременно и полезно было, то имеет оное само себя править» , том 50, №2
Аллергия | Симптомы, осложнения, диагностика и лечение
Аллергические реакции возникают, когда ваша иммунная система реагирует на чужеродное вещество, такое как пыльца, пчелиный яд, шерсть или пища, которая не вызывает реакции у большинства людей.
Иммунная система производит вещества, известные как антитела. Когда у вас аллергия, иммунная система создает антитела, которые идентифицируют конкретный аллерген как вредный, хотя это не так. Когда вы вступаете в контакт с аллергеном, реакция иммунной системы может воспламенить кожу, пазухи, дыхательные пути или пищеварительную систему.
Тяжесть аллергии варьируется от человека к человеку и может варьироваться от незначительного раздражения до анафилаксии — потенциально опасной для жизни чрезвычайной ситуации. Хотя большинство аллергии нельзя вылечить, лечение может помочь облегчить симптомы.
Симптомы аллергии, которые зависят от участвующего в процессе вещества, могут влиять на дыхательные пути, пазухи и носовые ходы, кожу и пищеварительную систему. Аллергические реакции могут варьироваться от легких до тяжелых. В некоторых серьезных случаях аллергия может вызвать опасную для жизни реакцию, известную как анафилаксия.
Аллергический ринит может вызвать:
- Чихание
- Зуд носа, глаз или верхней части рта
- Заложенность носа
- Красные или опухшие глаза (конъюнктивит)
Пищевая аллергия может вызвать:
- Покалывание во рту
- Отек губ, языка, лица или горла
- Крапивница
- Анафилаксия
Аллергия на укус насекомых может вызвать:
- Большая площадь набухания (отека) на месте укуса
- Зуд или воспаления по всему телу
- Кашель, сжатие грудной клетки, одышка
- Анафилаксия
Аллергия на лекарственные препараты может вызвать:
- Крапивница
- Зуд
- Высыпание
- Опухоль частей лица
- Анафилаксия
Атопический дерматит, аллергическое состояние кожи, также называемое экземой, может привести к:
- Зуд
- Покраснения
- Шелушение кожи
Анафилаксия
Некоторые виды аллергии, включая аллергии на продукты питания и укусы насекомых, могут вызвать серьезную реакцию, известную как анафилаксия. Анафилаксия может вызвать шок. Признаки и симптомы анафилаксии включают:
- Потеря сознания
- Падение артериального давления
- Тяжелая одышка
- Кожная сыпь
- Головокружение
- Быстрый/слабый пульс
- Тошнота и рвота
Аллергия начинается, когда иммунная система ошибочно воспринимает нормально безопасное вещество для опасного захватчика. Затем иммунная система вырабатывает антитела, которые остаются начеку для этого конкретного аллергена. Когда вы снова подвергаетесь воздействию аллергена, эти антитела могут выделять ряд химических реакций иммунной системы, таких как гистамин, которые вызывают симптомы аллергии.
Общие триггеры аллергии включают:
- Воздушные аллергены, такие как пыльца, шерсть животных, клещи и плесень
- Определенные продукты, в частности арахис, орехи, пшеница, соя, рыба, моллюски, яйца и молоко
- Укусы насекомых, например, от пчелы или осы
- Лекарства, особенно пенициллин или антибиотики на основе пенициллина
- Латекс или другие вещества, которые вы касаетесь, которые могут вызывать аллергические кожные реакции
У вас может быть больше шансов на развитие аллергии, если вы:
- Обладаете хронической астмов или аллергией
- Воспитываете ребенка
- Астматик или аллергик
Осложнения
Наличие аллергии увеличивает риск некоторых других медицинских проблем, в том числе:
- Анафилаксия.
 Если у вас серьезные аллергии, вы подвергаетесь повышенному риску этой серьезной реакции, вызванной аллергией. Продукты питания, лекарства и укусы насекомых являются наиболее распространенными пусковыми механизмами анафилаксии.
Если у вас серьезные аллергии, вы подвергаетесь повышенному риску этой серьезной реакции, вызванной аллергией. Продукты питания, лекарства и укусы насекомых являются наиболее распространенными пусковыми механизмами анафилаксии. - Астма. Если у вас аллергия, у вас, скорее всего, будет астма – реакция иммунной системы, которая влияет на дыхательные пути и дыхание. Во многих случаях астма вызвана воздействием аллергена в окружающей среде.
- Синусит и инфекции ушей или легких. Риск выше, если у вас есть астма.
Профилактика
Предотвращение аллергических реакций зависит от типа аллергии. Общие меры включают следующее:
- Избегайте известных триггеров. Даже если вы лечите симптомы аллергии, старайтесь избегать триггеров. Если, например, у вас аллергия на пыльцу, оставайтесь внутри, когда окна и двери закрыты, когда пыльца высока. Если у вас аллергия на пылевидные клещи, пыль и вакуум, меняйте постельные принадлежности часто.
- Вести дневник. При попытке определить, что вызывает или ухудшает аллергические симптомы, отслеживайте свои действия и то, что вы едите, когда появляются симптомы и что, по-видимому, помогает. Это может помочь вам и вашему врачу определить триггеры.
- Наденьте браслет медицинского предупреждения. Если у вас была сильная аллергическая реакция, медицинский браслет предупреждает других о том, что у вас серьезная аллергия, если у вас есть реакция, не позволяющая отвечать на вопросы окружающих людей.
Диагностика
Чтобы оценить, есть ли у вас аллергия, ваш врач, скорее всего:
- Задаст подробные вопросы о симптомах
- Выполнит физический экзамен
- Уточнит, ведете ли вы дневник симптомов или возможных триггеров
Если у вас есть пищевая аллергия, ваш врач, скорее всего:
- Попросит вас вести подробный дневник продуктов, которые вы едите
- Спросит, прекратили ли вы есть подозрительную пищу во время оценки аллергии
Ваш врач может также рекомендовать один или оба из следующих тестов. Однако имейте в виду, что эти тесты на аллергию могут быть ложно положительными или ложно отрицательными.
- Кожный тест. Врач или медсестра будут колоть кожу и тестировать на вас небольшое количество белков, обнаруженных в потенциальных аллергенах. Если у вас аллергия, у вас, вероятно, будет развиться повышенная реакция в месте тестирования на вашей коже.
- Анализ крови. Специфическое тестирование крови, обычно называемое тестированием с помощью радиоаллергербората (RAST) или ImmunoCAP, измеряет количество антител, вызывающих аллергию, в вашем кровотоке, известных как иммуноглобулиновые E (IgE) антитела. Образец крови отправляется в медицинскую лабораторию, где ее можно проверить на предмет наличия чувствительности к возможным аллергенам.
Если ваш врач подозревает, что ваши проблемы вызваны чем-то иным, чем аллергия, другие тесты могут помочь выявить или исключить другие медицинские проблемы.
Лечение аллергии включает:
- Изоляция от аллергенов. Ваш врач поможет предпринять шаги, чтобы выявить и избежать аллергию. Это, как правило, самый важный шаг в предотвращении аллергических реакций и уменьшении симптомов.
- Лекарственные препараты. В зависимости от аллергии, лекарства могут помочь уменьшить реакцию иммунной системы и облегчить симптомы. Ваш врач может предложить лекарство без рецепта или рецептурные лекарства в виде таблеток, назальных спреев или глазных капель.
- Иммунотерапия. Это лечение включает в себя ряд инъекций очищенных экстрактов аллергенов, обычно даваемых в течение нескольких лет.
Другой формой иммунотерапии является таблетка, которая помещается под язык (подъязычный) до тех пор, пока она не растворится. Подъязычные препараты используются для лечения некоторых аллергий на пыльцу.
- Экстренный эпинефрин. Если у вас тяжелая аллергия, вам может потребоваться срочная доза эпинефрина. При тяжелых аллергических реакциях, эпинефрин (Auvi-Q, EpiPen, другие) может уменьшить симптомы, пока вы не получите экстренное лечение.
У глаз есть это: почему природа (иногда) останавливается всего на двух | by Naturalish
Буквально на прошлой неделе ветеринар в Великобритании сделал шокирующее открытие: 4-месячная корова с третьим глазом шлепнулась по центру лба. «Я никогда раньше не видел ничего подобного», — сказал ветеринар, и многие из нас, читающих Интернет, должны были согласиться с этим, учитывая, как быстро эта история распространилась по сети.
Это жуткое, тревожное явление природы… или нет?
Как сообщает Wales’ Daily Post: «У него есть веки и ресницы, и он тоже влажный, как будто выделяется какая-то смазка». Спасибо за такие богатые подробности! Кажется, что два глаза — это естественный порядок вещей. Люди рождаются с двумя глазами, собаки и кошки рождаются с двумя глазами, и часто трудно представить себе, что может повлечь за собой альтернатива, хотя и не из области фантастики и фантазии. Многоглазые существа были основным продуктом мифологии на протяжении тысячелетий, поэтому очевидно, что мы, двуглазые люди с богатым воображением, способны мыслить за пределами наших биологических ограничений, по крайней мере, до некоторой степени.
А на самом деле? Это кажется слишком чуждым, по крайней мере, на первый взгляд — пока мы не заглянем глубже в богатое разнообразие мира природы и не поймем, что эволюция не предвзято относится к многоглазым среди нас — будь то три, восемь или далеко в сотни. То, что кажется сверхъестественным произведением чистой фантастики, на самом деле имеет глубокие прецеденты в реальной биологии. Во многих случаях это даже более странно, чем мы привыкли считать возможным.
Глаза: правда или вымысел?
Давайте сначала избавимся от моего риторического титула. Почему биология — по крайней мере, человек биология — предпочитает двуглазый шаблон?
Мы все были там.Это не ракетостроение, хотя математика может быть довольно сложной, если вы увлекаетесь подобными вещами. Бинокулярное (двуглазое) зрение дает нам возможность воспринимать глубину в дополнение к цвету и деталям, которые мы могли бы обрабатывать только через одну точку.
При использовании двух глаз играют роль два дополнительных фактора: один не очень важный, а именно акт конвергенции или дивергенции глаз, а другой очень важный, а именно стереоскопическое восприятие глубины в силу непохожесть изображений, представляемых трехмерным объектом или массивом объектов, для отдельных глаз.
Но зачем останавливаться на достигнутом?
Глаза являются дорогим органом — это означает, что «метаболические затраты» на глаза значительны — достаточно для того, чтобы эволюционный процесс (просто говоря) «оправдал» инвестиции только в том случае, если они будут перевешиваться выгодами для нашего благополучия во всем жизнь. Если бы мы не получали существенной пользы от наличия двух глаз, этого, вероятно, не произошло бы — и через некоторое время, если бы три глаза (или больше) были еще более ценными и перевешивали инвестиции нашего тела в более сложные органы, вполне вероятно, что мы бы видели, что адаптация возникает в нашей родословной.
Только по этой причине я ненавижу коз, но уважаю там, где это необходимо. Глаза у них небрежные, но они выполняют свою работу. «Два глаза» действительно являются наиболее оптимальной стратегией для нашего образа жизни — и это относится ко всем млекопитающим. Конечно, в классе еще много разнообразия. У приматов (как и у нас) два глаза направлены вперед для лучшего восприятия глубины, но более узкое поле зрения, около 120°. Другие виды млекопитающих, особенно те, на которых часто охотятся как на добычу (с козой в качестве крайней меры), имеют поля зрения более 320 ° с очень небольшим перекрытием.
А в еще более крайних случаях, зачем вообще глаза? Вопреки распространенному мнению, у млекопитающих, таких как крот-звездонос, есть глаза, но они сильно недоразвиты и связаны с мозгом «крошечными зрительными нервами», гораздо меньшими, чем можно было бы ожидать от существ их размера.
Это возвращает нас к нашей истории с коровами-мутантами с новой ценной информацией. Три глаза (а возможно, и больше) вполне возможны в эволюции млекопитающих, но даже если бы третий глаз теленка был полностью оборудован для сбора световых сигналов, он должен быть подключен к зрительной коре, способной обрабатывать эту информацию. В царстве млекопитающих всегда будут господствовать два глаза. Просто так мы устроены.
Но можно представить и альтернативы.
У меня есть вопросы о том, могли ли нибблонийцы эволюционировать среди флоры Вергона 6 или сходство — просто совпадение. Я признаю, что знания о Футураме в наши дни немного заржавели. Это были жаркие 22 года.Некоторые подобные творения научной фантастики красивы и прямолинейны. Нибблер из «Футурама » представляет нам трехглазое существо, которое может видеть поверх высокой травы планеты Вергон 6 или обеспечивать своего рода камуфляж среди листвы. То есть до тех пор, пока история нибблонианского происхождения не сойдет с рельсов.
Я считаю, что Beholder является культурным каноном, эквивалентным греческому мифу на данный момент — дайте мне знать, если это кого-то раздражает. Углубляясь в знания, мы натыкаемся на таких существ, как Аргус, многоглазый греческий великан (вверху слева) или архетипический злодей Dungeons & Dragons Смотрящий (вверху справа). Теперь мы куда-то движемся. Как это часто бывает в подобных многоглазых мифах, множество глаз наделяют исключительными впечатляющими способностями. Бехолдер может замедлять, окаменевать или разрушать врагов лучами, исходящими из каждого стебля глаза. Для Аргуса глаза даруют непоколебимое и всеведущее зрение — по некоторым сведениям, всевидящее и всезнающее.
Это образ в художественной литературе, который появляется снова и снова — множество глаз, которые дают зрение за пределами нашего обычного мира. Иногда именно «третий глаз» предлагает «восприятие за пределами обычного зрения». В христианской символике «множество «глаз» павлина на его хвосте указывало на всеведение Бога». Глядя в целом на совокупность художественной литературы и мифа, существа с большим количеством глаз, как правило, не просто видят лучше и глубже — они видят далеко за пределами человеческих возможностей. Они видят сквозь пространство, время или частную жизнь. Это линия, проведенная между нашим реальным, ограниченным, двуглазым миром и пределами воображения.
Верно?
Экстремальные глаза природы
Люди часто попадают в ловушки «млекопитающего» шаблона, который мы иногда используем, чтобы сформировать наше восприятие биологии в целом. По правде говоря, правила, которые диктуют нашу анатомию, и окончательные строительные блоки наших харизматичных, покрытых мехом, кормящих кузенов, серьезно ограничивают понимание самых реальных крайностей природы вокруг нас. Хотя приятно проводить тонкую грань между «два глаза = нормально» и «больше глаз = причудливо», мир определенно не так устроен.
И даже то «зрение за пределами видимости», которое мы любим относить к строгой фантастике и мифологии, является, откровенно говоря, реальным явлением природы, которое обычно не усваивается, когда мы думаем об эволюции.
Для начала теменный глаз .
Здесь изображена молодая американская лягушка-бык. Какая прелесть! У бесчисленного множества амфибий, акул, ящериц и знаменитой рептилии, не являющейся ящерицей, называемой туатара, теменной глаз является светочувствительным органом, расположенным на макушке головы. Обычно это находится прямо между двумя «нормальными» глазами — я знаю, что это постыдно ориентированная на млекопитающих фаза, которую мне, вероятно, следует избегать.
Хотя функциональное существование этого органа неодинаково для всех групп животных и часто сбивает с толку специалистов по поведению животных, некоторые исследования показывают, что чувствительность органа к свету можно откалибровать, чтобы он действовал как компас и позволял более точно ориентироваться по пути солнца. .
Настоящий «третий глаз», позволяющий животным видеть больше, чем видят люди? Эгад!
А членистоногих мы еще даже не обсуждали.
Хорошо, теперь мы кое-что получили.Не у всех пауков восемь глаз, но у тех, у кого они есть, эти дополнительные органы чувств наделяют удивительными способностями:
Дополнительные глаза выполняют множество функций. В некоторых случаях боковые глаза расширяют круг первичных глаз, давая паукообразному широкоугольное изображение. Вторичные глаза действуют как детекторы движения и предоставляют информацию о восприятии глубины, помогая пауку определить расстояние, а также направление добычи или угрозы.
И мы не можем забыть (снова) креветку-богомола! Эти плохие парни имеют до 16 разновидностей фоторецепторных клеток, по сравнению с ничтожными тремя у млекопитающих, и могут видеть цвета и поляризацию, которые люди никогда не смогли бы обнаружить естественным путем без усовершенствованного оборудования. Более того, они способны обрабатывать глубину каждого глаза без необходимости синтезировать несколько изображений в мозгу, подобно нам — это то, что ученые называют «тринокулярным зрением».
Скромный хитон: Морской Аргус! (©) Список продолжает включать все более и более экстремальные, умопомрачительные случаи захватывающего зрелища в глубинах животного мира. Последнее существо в нашем списке зарезервировано для хитона, маленького плоского моллюска, который часто посещает приливные зоны по всей планете и обладает сотнями крошечных глаз, разбросанных по их панцирям. В 2015 году дальнейшие исследования улучшили наше понимание зрения хитона и пришли к выводу, что глаза не так примитивны, как мы когда-то думали — на самом деле они способны формировать сфокусированные изображения и помогают хитону обнаруживать движение, идущее под почти угловым углом поверхности. .
Стоглазый Аргус или всеведущий «Третий глаз» из мифов кажутся гораздо менее причудливыми по сравнению со слайд-шоу крайностей природы.
Было бы ошибкой смотреть на мир — буквально — только глазами млекопитающих. Хотя нам хотелось бы думать, что наша точка зрения является нормой, на самом деле это далеко не так. На протяжении веков рассказчики постулировали о невозможных сценариях с помощью видения, способного к вещам, далеким от нашей анатомии, когда на самом деле природа уже придумала немало крайностей, которые превосходят даже наши самые смелые фантазии.
Все это говорит о том, что если вы корова в Уэльсе, которую дразнят из-за вашего третьего глаза, вы на самом деле в отличной компании — правда и вымысел.
Глаза млекопитающих раскрывают темное прошлое
На протяжении более 100 миллионов лет динозавры доминировали в светлое время суток, а солнечное тепло позволяло им процветать. Таким образом, ночь могла предоставить мелким, преимущественно насекомоядным млекопитающим, наилучшую возможность поохотиться, не становясь следующей едой динозавра.
Считается, что ранние млекопитающие, появившиеся в тени динозавров, вели в основном ночной образ жизни. У млекопитающих развилось острое обоняние и слух, и за счет некоторой способности видеть в цвете и с более высоким разрешением их глаза развили приспособления, которые улучшили их зрение в темноте: увеличенные зрачки, которые позволяют большему количеству света проникать в глаз; большее количество палочек, фоторецепторов, необходимых для зрения в тусклом свете; и отражающий слой ткани, называемый Tapetum lucidum, который увеличивает поглощение света сетчаткой.
Изучая глаза и гены современных животных, а также изучая летопись окаменелостей, ученые начали собирать воедино сведения о том, как долго млекопитающие были прикованы к ночи и как этот период сформировал зрительную систему современных млекопитающих. Такая работа также предлагает понимание того, почему человеческие глаза иногда колеблются. Хотя детали остаются предметом обсуждения, понимание эволюции зрения млекопитающих в течение продолжительного периода ночного образа жизни также может помочь ученым предсказать, как современные млекопитающие могут адаптироваться к растущему световому загрязнению и деятельности человека, заставляющей некоторых дневных животных восстанавливать ночное время.
«То, что мы имеем сегодня с млекопитающими, — это излучение организмов, которые развили наиболее существенные черты своих сенсорных систем в ночных условиях», — говорит Кристофер Кирк, биологический антрополог из Техасского университета в Остине. «Сто миллионов лет ночной жизни — я уже говорил это раньше и скажу снова — у этого есть последствия».
Рождение гипотезы
В своей монографии 1942 года « Глаз позвоночных и его адаптивное излучение » Гордон Уоллс, биолог из тогдашнего Университета Уэйна в Детройте, штат Мичиган, предположил, что во время ранней эволюции млекопитающих, чтобы объяснить, почему глаза млекопитающих отличаются от глаз других позвоночных. Эта идея стала известна как гипотеза «ночного узкого места».
Многие странности млекопитающих можно объяснить тем, что они ведут ночной образ жизни. Помимо больших роговиц и зрачков, чтобы максимизировать количество света, которое может попасть в глаз, глаза млекопитающих имеют меньшее расстояние между хрусталиком и сетчаткой, чем глаза многих позвоночных, что помогает хрусталику проецировать яркое изображение на сетчатку. в тусклом свете. Существует также меньше типов фоторецепторных клеток для обнаружения цветов (известных как колбочки). А в глазах большинства млекопитающих — хотя в особенности не у людей или некоторых других приматов — отсутствует ямка, область сетчатки, богатая колбочковыми клетками, которая обеспечивает четкое и детальное зрение рыбам, птицам и рептилиям, охотящимся днем. «Глаза большинства млекопитающих не так хороши, как у птиц, ящериц и даже рифовых рыб», — говорит Ларс Шмитц, биолог-эволюционист из колледжа Клермонт Маккенна в Калифорнии.
В последнее десятилетие исследователи использовали новые инструменты и подходы для сбора данных, чтобы поддержать гипотезу ночного узкого места. В 2012 году Кирк и его коллеги проанализировали морфологию глаз 266 видов млекопитающих из 23 отрядов 1 . Они обнаружили, что форма практически не различалась, независимо от того, были ли виды активны днем, ночью или и тем, и другим. Это отличало млекопитающих от птиц и ящериц, форма глаз которых больше различается у ночных и дневных видов. Глаза млекопитающих, как правило, напоминали глаза ночных птиц и ящериц, за исключением людей и близкородственных обезьян и человекообразных обезьян. Кирк предполагает, что это связано с тем, что глаза некоторых млекопитающих, вероятно, начали «переэволюционировать» черты, полезные для дневной жизни, после того, как они отказались от ночного образа жизни.
Исследователи выясняют, когда наступил этот период ночной жизни и как долго он длился. В 2017 году группа из США предположила, что предки млекопитающих вели дневной образ жизни. Они использовали данные окаменелостей, чтобы показать, что около 385 миллионов лет назад (как раз перед тем, как позвоночные вышли из воды на сушу 2 ) глаза позвоночных утроились в размере и переместились с боков на макушку головы. Эти приспособления позволили ранним позвоночным видеть намного дальше и охотиться, глядя над водой, как это делают крокодилы. Это было бы наиболее выгодно при ярком свете, говорит Рой Маор, эколог млекопитающих из Тель-Авивского университета в Израиле и Университетского колледжа Лондона, предполагая, что ранние позвоночные были активны днем.
Что-то должно было измениться, чтобы млекопитающие стали вести ночной образ жизни. В конце 2017 года группа исследователей, в том числе Маор, представила доказательства того, что это может быть. Команда использовала данные о моделях ежедневной активности 2415 живых видов млекопитающих, чтобы реконструировать два потенциальных генеалогических дерева — одно охватывает 166 миллионов лет истории эволюции млекопитающих, а другое — 218 миллионов лет 3 . Дерево с более короткой временной шкалой предполагает, что млекопитающие начали занимать дневную нишу после того, как господство нептичьих динозавров закончилось вымиранием мелового/палеогенового периода, вызванным падением астероида на полуостров Юкатан на территории современной Мексики, около 66 миллионов лет назад. назад. Это исследование было первым, в котором модели активности млекопитающих были привязаны к четкой временной шкале, говорит Маор, и подтверждает гипотезу о том, что ранние млекопитающие были вынуждены вести ночной образ жизни из-за преобладания динозаврами дневных часов.
Меловое/палеогеновое вымирание «было лучшим событием для млекопитающих», — говорит Кирк. «Без этого 10-километрового куска скалы, врезающегося в Юкатан, кто знает, как бы выглядели современные млекопитающие — вероятно, они все еще были бы просто кучей маленьких землероек, пытающихся не стать обедом».
Серая зона
Однако не все данные подтверждают эту версию гипотезы ночного узкого места. Некоторые исследования, например, намекают на то, что время бодрствования млекопитающих и динозавров могло иметь большее совпадение, чем предполагает гипотеза.
Например, есть свидетельства того, что у ранних млекопитающих были варианты генов, которые кодируют светочувствительные белки, называемые опсинами, которые настроены на цветовое зрение. В 2012 году эти данные были использованы для предположения, что активность млекопитающих могла не ограничиваться темными часами 4 . Комбинация опсинов у таких млекопитающих напоминала таковую у современных змей, активных при тусклом свете. Таким образом, млекопитающие могли охотиться в сумерках, а не вести полностью ночной образ жизни.
Некоторые динозавры, вероятно, также выходили на улицу после захода солнца, говорит Шмитц. Вместе с Рёске Мотани, палеобиологом из Калифорнийского университета в Дэвисе, он изучил окаменелости 33 архозавров, группы, в которую входят динозавры и летающие рептилии, известные как птерозавры. Пара сосредоточила свой анализ на глазницах (орбитах) животных и склеральных кольцах 5 , которые представляют собой кости, окружающие глаза многих позвоночных.
Они начали со сравнения размеров склеральных колец с соответствующими им орбитами у более чем 150 современных животных и сопоставления полученных соотношений с моделями активности животных. Затем они использовали эту информацию, чтобы сделать вывод об активности динозавров по размеру их глаз, что привело их к предположению, что некоторые динозавры действительно бродили в темноте. «Большинство динозавров были активны днем, а большинство млекопитающих — ночью», — говорит Шмитц. — Но, наверное, не ясно. Такое совпадение может помочь объяснить появление около 130 миллионов лет назад млекопитающего длиной почти 1 метр, питающегося динозаврами.
Есть также доказательства того, что не млекопитающие уходят в ночь, а их предки, возможно, сделали это, говорит Шмитц. В 2014 году он и Кеннет Ангельчик, палеобиолог из Полевого музея естественной истории в Чикаго, штат Иллинойс, проанализировали орбиты и склеральные кольца 24 видов синапсид немлекопитающих — млекопитающих рептилий, которые предшествовали настоящим млекопитающим. Экстраполируя форму глаз по размерам окаменелостей, пара обнаружила свидетельства ночного образа жизни, существовавшего более 300 миллионов лет назад — более чем за 100 миллионов лет до появления млекопитающих. 0101 6 . «Возможно, они перешли в ночную фазу даже раньше, чем мы думали», — говорит Шмитц.
Но Кирк и несколько других исследователей, в том числе Маргарет Холл, биолог-эволюционист и анатом из Университета Среднего Запада в Глендейле, штат Аризона, задались вопросом, дают ли размеры орбит и склеральных колец у современных животных достаточно информации для реконструкции поведения окаменелых существ. Холл и Кирк предполагают, что для предсказания того, как видят животные, требуется знание размера роговицы и диаметра глазного яблока, измерения, которые являются неопределенными для давно вымерших животных. «История, которую мы все рассказываем себе и друг другу, — что динозавры каким-то образом вынудили млекопитающих вести ночной образ жизни — может быть правдой», — говорит Холл. «Это самая правдоподобная история, которую мы можем придумать. Но узнать это невозможно».
Возвращение во тьму
Некоторые исследователи считают, что ночное узкое место может быть корнем некоторых проблем, влияющих на зрение человека. Например, приспосабливаясь к ночному образу жизни, млекопитающие, по-видимому, утратили механизм, с помощью которого современные ящерицы и птицы сжимают линзы, чтобы сфокусироваться. Это делает людей более уязвимыми к пресбиопии или дальнозоркости с возрастом. Точно так же возрастная дегенерация желтого пятна, при которой деградирует небольшая область сетчатки, что приводит к потере центрального зрения, имеет такой серьезный эффект из-за того, как люди и другие приматы приобрели остроту зрения: путем втисания колбочек в сетчатку. ямка. Эта структура, вероятно, возникла как приспособление к развитию сети кровеносных сосудов над основанием сетчатки после ночного узкого места. По словам Кирка, чтобы эта сеть не мешала острому зрению, клетки колбочек были вынуждены занимать небольшую — и, следовательно, уязвимую — площадь. «Когда вы спрашиваете, почему мы вообще должны беспокоиться об этих болезненных процессах, во многом это связано с утратой функциональности в узком месте и всем, что произошло, чтобы компенсировать это», — говорит он.
Современные млекопитающие, такие как европейский бобр ( Castor fiber ), часто активны ночью в городских районах. Предоставлено: Geslin Laurent
Этот процесс компенсации дает представление о том, как животные могут адаптироваться к жизни в среде, которая становится более яркой и более густонаселенной людьми. В метаанализе 2018 года исследователи в США изучили 76 исследований активности 62 видов млекопитающих на 6 континентах 7 . Млекопитающие, которые жили в районах с высоким уровнем беспокойства человека (или в более спокойных районах во время повышенной активности человека, например, в сезон охоты), вели ночной образ жизни на 36% больше, чем те, которые жили относительно спокойно.
Кейтлин Гейнор, эколог дикой природы из Калифорнийского университета в Беркли, которая работала над метаанализом, говорит, что исследователям трудно предсказать все экологические последствия смены ниши животными. В горах Санта-Крус в Калифорнии койоты, живущие рядом с популярными местами для пеших прогулок, стали вести более ночной образ жизни, и, в свою очередь, в их рацион теперь чаще входят животные, активные ночью. Это будет иметь последствия для их новой добычи, а также для ночных хищников, включая лисиц, с которыми они сейчас конкурируют. И неясно, насколько успешно другие дневные млекопитающие будут общаться, питаться и избегать хищников после перехода на ночной образ жизни.
По мере того, как уровень человеческой активности продолжает увеличиваться, люди могут создать еще одно узкое место в ночное время. «Млекопитающие наслаждались солнечным светом с тех пор, как вымерли динозавры, но теперь мы загоняем их обратно в ночное время», — говорит Гейнор. «Мы — вездесущая, ужасающая сила на планете — так же, как динозавры».
Ссылки
Hall, M.I., Kamilar, J.M. & Kirk, E.C. Proc. биол. науч. 279 , 4962–4968 (2012).
ПабМед Статья Google ученый
Макивер, М. А., Шмитц, Л., Муган, У., Мерфи, Т. Д. и Мобли, К.
Д. Proc. Натл акад. науч. США 114 , E2375–E2384 (2017).ПабМед Статья Google ученый
Маор Э., Даян Т., Фергюсон-Гоу Х. и Джонс К. Э. Nature Ecol. Эвол. 1, 1889–1895 (2017).
ПабМед Статья Google ученый
Дэвис В.И., Коллин С.П. и Хант Д.М. Мол. Экол. 21, 3121–3158 (2012).
ПабМед Статья Google ученый
Schmitz, L. & Motani, R. Science 332 , 705–708 (2011).
ПабМед Статья Google ученый
Angielczyk, K.D. & Schmitz, L. Proc. биологических наук. 281 , 20141642 (2014).
ПабМед Статья Google ученый
Гейнор, К.М., Хойновски, К.Е., Картер, Н.Х. и Брашарес, Дж.С. Science 360 , 1232–1235 (2018).
ПабМед Статья Google ученый
Скачать ссылки
У этого вымершего варана было четыре глаза
- Животные
Четыре глаза лучше, чем два, по крайней мере, так было у варанов, живших около 50 миллионов лет назад.
Автор Jen Viegas
Опубликовано 02.04.2018 в 15:23
Окаменелость Saniwa ensidens в Полевом музее естественной истории | Wikimedia Commons
Окаменелость Saniwa ensidens в Полевом музее естественной истории | Викисклад
Почти все животные имеют два глаза, но есть редкие исключения. Например, ракообразные семейства Cyclopidae, также известные как Cyclops, видят мир одним большим глазом. На другой стороне спектра находятся гигантские моллюски, у которых сотни глаз. Среди позвоночных рекордсменом является бесчелюстная минога. Эта рыба, напоминающая угря, принадлежит к очень древнему роду и обладает четырьмя глазами.
Дополнительные два глаза представляют собой фотосенсорные структуры на макушке головы, известные как шишковидный и парапинеальный органы. Ученые все еще пытаются определить все функции этих органов, но известно, что они играют ключевую роль в ориентации и в циркадных и годовых циклах.
Пинеальный орган обнаружен у ряда низших позвоночных, таких как рыбы и лягушки, но парапинеальный орган встречается реже. Кристер Смит из Научно-исследовательского института Зенкенберга, однако, давно подозревал, что древние ящерицы могли иметь обе фотосенсорные структуры вместе с еще двумя типичными глазами.
«Идея впервые зародилась, когда я, будучи аспирантом, изучал разнообразие ископаемых ящериц в коллекциях Йельского университета, — сказал Смит. «В одном случае были противоречивые интерпретации третьего глаза в теменной кости разными экспертами. Было ясно, что в кости было два больших отверстия, одно за другим, и я вспомнил, что было два шишковидных органа».
Смит признает, что искать древнюю четырехглазую ящерицу было «дурацкой» идеей, но она и ее команда так и сделали.
Они исследовали музейные экспонаты, собранные почти 150 лет назад в Гризли-Баттс, месте в бассейне Бриджер в штате Вайоминг. Компьютерная томография показала, что у двух разных особей вымершего вида варанов (Saniwa ensidens) были промежутки на месте двух дополнительных глаз.
Открытие, опубликованное в журнале Current Biology , документирует первое известное четырехглазое челюстное позвоночное.
«У ученого много идей, но многие — даже большинство из них — не реализуются, и чем оригинальнее идея, тем выше вероятность ее провала», — сказала Смит, добавив, что она была взволнована, когда ее подозрения относительно окаменелых ящериц подтвердились.
Открытие проливает свет на эволюцию дополнительных глаз.
На этом изображении представлена реконструкция того, как мог выглядеть вымерший варан. На наложенном черепе видны теменное и шишковидное отверстия. | Senckenberg Gesellschaft für Naturforschung/Andreas Lachmann/Digimorph.orgСмит и ее коллеги Бхарт-Анджан Бхуллар, Гюнтер Келер и Йорг Хаберсетцер подозревают, что у миног развились четыре глаза одновременно, но они говорят, что потребуются дополнительные находки окаменелостей, чтобы определить, когда появился каждый из органов.
Известно, что у самых ранних млекопитающих — наших далеких предков — было не просто два глаза.
«У стволовых млекопитающих было три глаза, — объяснил Смит. «Было высказано предположение, что ночная фаза эволюции предков млекопитающих привела к исчезновению третьего глаза в этой линии, и, возможно, что-то подобное произошло в других группах четвероногих».
Таким образом, многие четвероногие или четвероногие животные могли потерять свой третий глаз, когда некоторые из них стали активными ночью и, вероятно, не нуждались в дополнительных фотосенсорных органах.
У людей и большинства других млекопитающих не осталось следов третьего глаза.
«В частности, у людей головной мозг настолько сильно расширен, что он плотно и полностью покрывает шишковидный орган, что делает практически невозможным для фотонов его достижение», — сказал Смит. «Кроме того, парапинеальный орган считается полностью отсутствующим».
СВЯЗАННЫЙ: Улучшенное зрение могло привлечь рыбу на сушу миллионы лет назад
Однако у крыс шишковидная железа находится близко к поверхности. Некоторые светочувствительные белки все еще экспрессируются в органе у эмбрионов крыс, что позволяет предположить, что третий глаз не полностью исчез у грызунов.
Смит и ее команда не уверены, какой орган — шишковидный или парапинеальный — привел к формированию третьего глаза у ранних млекопитающих. Когда эта загадка будет раскрыта, они должны лучше понять, почему млекопитающие со временем потеряли дополнительный глаз.
Что более ясно, так это то, что третий глаз современных ящериц, шишковидная железа, развился независимо от третьего глаза других челюстных позвоночных.
«Мы должны подчеркнуть, что идея о том, что третий глаз ящерицы — это не то же самое, что третий глаз лягушки или рыбы, высказывалась ранее», — сказал Смит. «Но это не было общепринятым. Обнаружив вид, у которого оба шишковидных органа были одновременно развиты как глаза, где передний был явно типичным третьим глазом ящерицы, мы смогли подтвердить эту идею».
Хотя дополнительные глаза вымершего варана располагались близко друг к другу, исследователи не думают, что они функционировали как пара так же, как обычные глаза.
«Они действительно расположены близко друг к другу, но их центральные нейронные связи с остальным мозгом другие, — объяснил Смит.
Saniwa ensidens жили в некоторых частях Вайоминга и Европы в эпоху эоцена около 48 миллионов лет назад. Он считается первой ископаемой ящерицей, когда-либо известной из Северной Америки. Теперь у него есть еще одна претензия на славу животных — по крайней мере, на данный момент.
Новое открытие «говорит нам, как легко, с точки зрения эволюции, самостоятельно собрать сложный орган при определенных обстоятельствах», — сказал Бхуллар, палеонтолог из Йельского университета. «Глаза классически рассматриваются как эти удивительно сложные структуры. На самом деле, мозг всегда только и ждет, чтобы сделать глаза».
Он добавил, что глаза являются «фундаментальной частью мозга» и что их формирование «является частью процесса контакта мозга с частью кожи». 0292 Смит сказал, что по мере развития эмбриона центральная нервная система начинается как «складывание самого внешнего слоя эмбриона». Этот слой называется нейроэктодермой.
СВЯЗАННЫЕ: генная терапия показала ранний успех в лечении общей причины слепоты Он содержит светочувствительный пигмент родопсин, присутствующий в глазах. Более недавнее исследование показало, что меняющий цвет боров также может видеть своей кожей. Возможно, что у других животных, в том числе у людей, кожа тоже обладает светочувствительными свойствами.
На данный момент исследователи больше сосредоточены на изучении ящериц. Смит и Бхуллар объяснили, что так называемый «сдвиг ящериц» — когда у ящериц впервые появился третий глаз — связан с развитием зрения у ряда различных позвоночных. Ученые надеются выяснить, когда произошло это важное изменение.
Смит сказала, что она и ее коллеги с нетерпением ждут возможности снова проанализировать коллекцию окаменелостей Гризли Баттс, несмотря на то, что останки «весьма обрывочны и потрепаны». Она пришла к выводу, что такие исследования «подчеркивают решающую роль, которую играют музеи как архивы биоразнообразия, как прошлого, так и настоящего».
Исследование говорит, что глаза эволюционировали для рентгеновского зрения
28 августа 2008 г.
У большинства животных глаза обращены вбок которые позволяют панорамный вид почти на все, что вокруг них, как спереди, так и сзади. Глаза некоторых млекопитающих эволюционировали до указывают в том же направлении. В то время как животные с вперед обращенные глаза теряют способность видеть то, что находится позади них, они приобретают рентгеновское зрение, что позволяет им видеть сквозь беспорядок в мире. |
Глаза, обращенные вперед, позволяют животным «видеть насквозь». беспорядок в мире
Преимущество использования двух глаз, чтобы видеть мир вокруг нас уже давно ассоциируется исключительно с нашей способностью видеть в 3D. А теперь новое исследование ученого из Rensselaer. Политехнический институт обнаружил поистине поразительную Преимущество бинокулярного зрения: наша способность видеть сквозь вещи.
Большинство животных — рыбы, насекомые, рептилии, птицы, кролики и лошади, например, существуют в незахламленной среде, такой как поля или равнины, а глаза у них расположены по обе стороны от их голова. Эти обращенные вбок глаза позволяют животному видеть перед собой и позади себя, способность, также известная как панорамное зрение.
Люди и другие крупные млекопитающие — приматы и крупные
плотоядные, такие как тигры, например, существуют в захламленных
такие среды, как леса или джунгли, и их глаза имеют
эволюционировали, чтобы указывать в том же направлении. В то время как животные с
глаза, обращенные вперед, теряют способность видеть то, что находится позади них,
они приобретают рентгеновское зрение, по словам Марка Чангизи, ассистента
профессор когнитивных наук в Ренсселере, который говорит глаза
обращенные в одном направлении, были выбраны для максимизации нашего
способность видеть в лиственной среде, такой как лес.
Все животные имеют область бинокля — части света что оба глаза могут видеть одновременно, что позволяет проводить рентгеновское зрение и растет по мере того, как глаза становятся более обращенными вперед.
Продемонстрировать наши рентгеновские способности довольно просто: возьмите ручку вертикально и смотреть на что-то далеко за его пределами. Если вы сначала закрой один глаз, а потом другой, ты увидишь, что в каждом случае ручка блокирует обзор. Однако, если вы откроете оба глаза, вы может видеть через перо мир за ним.
Чтобы продемонстрировать, как наши глаза позволяют нам видеть сквозь беспорядок, поднимите все пальцы в произвольном направлении и обратите внимание, как большую часть мира вы можете увидеть за ними, когда только один глаз открытый по сравнению с обоими. Вы многое упускаете одним глазом открытым, но может видеть почти все за беспорядком с оба.
«Наша бинокулярная область — своего рода «прожектор», светящийся
через беспорядок, позволяя нам визуально выметать
загроможденной области, чтобы распознавать объекты за ней», — говорит
Чангизи, главного исследователя проекта. «В качестве
пока расстояние между нашими глазами шире, чем ширина
объектов, вызывающих беспорядок — как в случае с нашим
пальцы, или было бы дело с листьями в лесу —
тогда мы сможем видеть сквозь него».
Чтобы определить, какие животные обладают такой впечатляющей силой, Чангизи изучил 319 видов 17 отрядов млекопитающих и обнаружил, что положение глаз зависит от двух переменных: беспорядок или его отсутствие в среде обитания животного, а также размер тела животного по отношению к объектам, создающим беспорядок.
Чангизи обнаружил, что животные в незахламленных средах, которые он описал как «нелистовые окружение или окружение, где находятся загромождающие объекты. больше по размеру, чем расстояние между глазами животного». (представьте, что крошечная мышь пытается видеть сквозь листья шириной 6 дюймов в в лесу) — глаза, как правило, обращены вбок.
«Животные за пределами листвы не должны иметь дело
с беспорядком, независимо от того, насколько они велики или малы, так что есть
у них никогда не было рентгеновского преимущества перед глазами, обращенными вперед». говорит Чангизи. «Потому что бинокулярное зрение не помогает им видеть
лучше, чем монокулярное зрение, они могут обозревать
гораздо большая область с повернутыми в сторону глазами».
Однако в загроможденной среде, которую определил Чангизи как лиственное окружение, где загромождающие объекты меньше чем расстояние между глазами животного — животные склонны имеют широкое поле бинокулярного зрения и, следовательно, смотрят вперед глаза, чтобы видеть сквозь листовые стены.
«Рентгеновское зрение позволяет животным с обращенные вперед глаза для визуального обзора гораздо большей области вокруг себя, чем позволили бы косые глаза», — говорит Чангизи. «Кроме того, чем крупнее животное в загроможденном окружающей среды, тем больше его глаза будут обращены вперед, чтобы позволить для максимально возможного рентгеновского зрения, чтобы помочь в охотиться, убегать от хищников и маневрировать в густых лес или джунгли».
Чангизи говорит, что человеческие глаза эволюционировали так, чтобы смотреть вперед,
но что теперь мы живем в незагроможденной среде, где мы
на самом деле может получить больше пользы от взгляда вбок.
«В современном мире у людей больше общего визуально с крошечные мыши в лесу, чем с большим животным в джунглях. Мы не сталкиваемся с большим количеством мелкого беспорядка, и вещи, которые загромождают наше поле зрения — автомобили и небоскребы — намного шире, чем расстояние между нашими глазами, поэтому мы мы не можем использовать наши рентгеновские лучи, чтобы видеть сквозь них», — говорит Чангизи. «Если мы заморозим себя сегодня и проснемся через миллион лет от теперь, возможно, нам будет трудно искать новую человеческую популяцию в глазах, потому что к тому времени они могли бы стоять боком».
Исследование Чангизи было завершено в сотрудничестве с Шинсуке Шимодзё из Калифорнийского технологического института и опубликовано в Интернете в Journal of Theoretical Биология. Он был профинансирован Национальными институтами Здоровье.
Исследования Чангизи в области рентгеновского зрения вместе с его исследованиями
о нашем
способность видеть будущее, цветовая телепатия и зрение
вычислительные способности, появится в его книге The
Vision Revolution (BenBella Books), поступит в продажу в этом
весна.
Контактное лицо: Эмбер Кливленд
Телефон: (518) 276-2146
Электронная почта: [email protected]
Внутривидовая изменчивость цвета глаз у птиц и млекопитающих: недавнее эволюционное событие, характерное исключительно для человека и домашних животных | Границы в зоологии
- Гипотеза
- Открытый доступ
- Опубликовано:
- Хуан Х. Негро 1 ,
- М. Кармен Блазкес 1,2 и
- Исмаэль Гальван 1
Границы зоологии том 14 , номер статьи: 53 (2017) Процитировать эту статью
7045 доступов
15 цитирований
72 Альтметрика
Сведения о показателях
Abstract
Background
Человеческие популяции и породы домашних животных состоят из особей с разнообразной окраской глаз (= радужной оболочки). Некоторые дикие птицы и млекопитающие могут иметь внутривидовую изменчивость цвета глаз, но эта вариация, по-видимому, связана со стадией развития особи, ее статусом размножения и/или половым диморфизмом. Другими словами, цвет глаз, как правило, является видоспецифическим признаком диких животных, и исключения составляют виды, у которых у особей одной возрастной группы или пола развивается одинаковый цвет глаз. Домашние животные, по определению, включают виды птиц и млекопитающих, искусственно отобранных человеком за последние несколько тысяч лет. Сами люди, возможно, приобрели разнообразную палитру цветов глаз также в недавнее эволюционное время, в мезолите или в верхнем палеолите.
Представление гипотезы
Мы выдвигаем две ранее непризнанные гипотезы относительно изменения цвета глаз: 1) окраска глаз у диких животных каждого вида имеет тенденцию быть фиксированным признаком. 2) У людей и популяций домашних животных, наоборот, глаза разноцветные. Половой отбор использовался для изменения цвета глаз у людей, но этот метод отбора не так легко применим к домашним животным, где спаривания контролируются заводчиком-человеком.
Проверка гипотезы
Окраска глаз у человека полигенна. Мы хотим исследовать генетику цвета глаз у других животных, а также экологические корреляты.
Последствия гипотезы
Изучение происхождения и функции цвета глаз прольет свет на причину, по которой у некоторых видов радужная оболочка может быть либо светлой (например, белой, желтой или светло-голубой), либо темной (темно-красной, коричневой). или черный). Причины, лежащие в основе огромного разнообразия цветов глаз у разных таксонов, никогда тщательно не исследовались, но вполне возможно, что работают все процессы дарвиновского отбора: половой отбор у людей, искусственный отбор у домашних животных и естественный отбор (в основном) у диких животных. животные.
Предыстория
Окраска животных привлекала экологов-эволюционистов с тех пор, как Дарвин и Уоллес спорили о вкладе естественного и полового отбора в эволюцию декоративных признаков. Если рассматривать позвоночных в целом, то цвет глаз и, в частности, цвет радужной оболочки действительно является изменчивым и бросающимся в глаза признаком [1], охватывающим практически все цвета радуги [2], будучи у некоторых видов ярко-желтыми или малиново-красными и у других видов очень темный и почти неотличимый от центрального зрачка, который всегда черный. Поэтому кажется, что некоторые цвета глаз могут способствовать рекламе, в то время как другие цвета менее заметны на лице или голове и могут служить для сокрытия глаза [3]. Окраска глаз также может быть связана со зрительными потребностями, поскольку задействованные пигменты улавливают разные длины волн света [4]. Тем не менее происхождение и функции цвета глаз до сих пор плохо изучены [5].
Представление гипотезы
Мы хотим сообщить о фундаментальном наблюдении, что изменчивость цвета радужной оболочки в основном межвидовая и что на внутривидовом уровне животные из классов птиц и млекопитающих имеют тенденцию иметь инвариантную видоспецифичную окраску. Это упущенное из виду правило нарушается людьми и домашними животными (как в классе птиц, так и в классе млекопитающих), которые демонстрируют изменчивость цвета глаз на уровне вида (рис. 1). Таким образом, у нас есть две рабочие гипотезы: 1) окраска глаз у диких животных имеет тенденцию быть постоянным признаком внутри вида, за исключением изменения цвета глаз во время взросления или связанного с половым дихроматизмом или статусом размножения. 2) У человека и домашних животных, наоборот, на видовом уровне глаза разноцветны. Мы будем утверждать, что эта вариация у людей и наших домашних животных возникла недавно в ходе эволюции. Это очевидно для домашних животных, чьи предки подверглись искусственному отбору в мезолите или неолите менее 10 000 лет назад. Но это также может быть верно и для людей, поскольку глаза светлого цвета впервые появились где-то до 8000 лет, возможно, в Северной Европе.
Цвет радужной оболочки у человека непрерывно варьируется от очень светло-голубого до темно-коричневого (верхняя линия, a , b , c , d ). Внутрипопуляционная изменчивость окраски глаз характерна и для домашних животных (средняя линия): двух взрослых кошек Felis catus ( e , f ) и двух взрослых домашних мускусных уток Cairina moschata domestica ( г , 7 ч ). Однако у диких животных цвет радужной оболочки, как правило, является фиксированным признаком с небольшими наблюдаемыми вариациями из-за взросления с возрастом или полового дихроматизма. В нижней строке показаны различия в окраске двух хищных птиц. Первый случай изменчивости, связанной с возрастом, у чернокрылых коршунов Elanus caeruleus ( i : взрослые особи; j : молодые особи). Во-вторых, вариации, связанные с полом: перепелятник Accipiter nisus ( k : взрослый самец; l : взрослая самка). и : CC BY-SA 2.0 (https://creativecommons.org/licenses/by-sa/2.0/, Luisangel https://flic.kr/p/4vHKkh). b : CC BY 2.0 (https://creativecommons.org/licenses/by/2.0/, великолепные глаза https://flic. kr/p/7vXh7G). c : CC BY 2.0 (https://creativecommons.org/licenses/by/2.0/, Жан-Симон Асселин https://flic.kr/p/2p4pFU). d : CC BY-SA 2.0 (https://creativecommons.org/licenses/by-sa/2.0/, Эмилио Кюффер https://flic.kr/p/agQvbv). и : CC BY 2.0 (https://creativecommons.org/licenses/by/2.0/, Триш Хамм https://flic.kr/p/dzfp8N). f : CC BY 2.0 (https://creativecommons.org/licenses/by/2.0/, Триш Хамм https://flic.kr/p/e8v8ro). k : CC BY 2.0 (https://creativecommons.org/licenses/by/2.0/, Энди Морффью https://flic.kr/p/KjN82N). l : CC BY 2.0 (https://creativecommons.org/licenses/by/2.0/, sighmanb https://flic.kr/p/ecc15G
Изображение в полный размер
Генетический вариант голубых глаз был извлечен из ДНК в останках охотников-собирателей на стоянке Мотала в Швеции и Лошбуре в Люксембурге, датированной 8000 лет до н.э. [6,7,8], а также в Мезолитические европейцы, жившие на Пиренейском полуострове 7000 лет назад [9]. Все эти данные, основанные на ДНК, позволяют предположить, что разноцветные глаза появились у Homo sapiens после их экспансии на запад в Европе [10], где они заменили неандертальцев около 40 000 лет назад [11]. В случае с домашними животными варианты со светлыми глазами обязательно закрепились в неолите или, возможно, когда-то раньше у собак (9).0095 Canis lupus familiaris ), первое в истории одомашненное животное [12]. Мы точно знаем, что существует различие в цвете глаз у тех видов, у которых дикие предки сохранились и имеют инвариантный цвет глаз, как у кошек и свиней. Однако мы должны признать, что у тех видов, предки которых уже вымерли, включая крупный рогатый скот и лошадей, мы можем отследить изменение цвета глаз с течением времени, только извлекая древнюю ДНК из останков.
Проверка гипотезы
Экологические корреляты окраски глаз
В скудной литературе по окраске глаз позвоночных межвидовая изменчивость приписывается не только отбору естественной средой, но также давлению хищников и половому отбору [5]. Исследование более 250 видов лягушек на Мадагаскаре показало, что ярко окрашенные радужки связаны с древесными лягушками, живущими в пологе леса, тогда как виды с темными глазами, как правило, обитают на земле [13]. Нам неизвестны сравнительные исследования окраски глаз млекопитающих, рептилий или рыб. Насколько нам известно, у птиц было опубликовано только два сравнительных исследования, оба на воробьиных. Один из них [14] сообщил о необъяснимо более высокой доле видов птиц с яркими радужками в Южной Африке (25%) и Австралии (35%) по сравнению с Канадой (6%) и Европой (8%). В другом исследовании [5] сообщалось, что птицы, гнездящиеся вне полостей, подвергаются жесткому отбору, чтобы развить темные глаза, и что цвет глаз не был связан с родительской заботой. На общем уровне также существует предположение, что преследующие хищники, будь то млекопитающие или птицы, обычно имеют желтые или светлые радужки, тогда как хищники, которые бегут за своей добычей, и сами жертвы, как правило, темноглазые. 2]. Несмотря на ограниченное количество исследований, в которых явно рассматривается изменение цвета глаз, все они предполагают, что отбор ограничивает изменчивость внутри вида.
Что касается внутривидовой изменчивости цвета радужной оболочки, признанные факторы изменения у птиц включают взросление с возрастом, изменения во время брачного сезона по сравнению с периодом отсутствия размножения, половой дихроматизм и подвидовую дифференциацию [15]. Что касается возрастных изменений окраски глаз, то для диких млекопитающих информация отсутствует, хотя хорошо известно, что у младенцев человека цвет глаз меняется в первые несколько месяцев жизни, а достижение окончательного цвета глаз может занять еще больше времени. У диких птиц возрастные изменения относительно широко распространены [16] и обычно затрагивают виды с ярко окрашенными радужными оболочками во взрослом возрасте, которые, однако, имеют тусклый цвет глаз в молодости [17]. Половой диморфизм в окраске глаз у птиц встречается редко [5] и неизвестен у млекопитающих, за исключением зеленых глаз у человека, которые чаще встречаются у женщин, чем у мужчин [18,19]. ,20].
Мы провели исследование внутривидовых изменений окраски у птиц в связи с взрослением с возрастом, половым диморфизмом или статусом размножения (дополнительные файлы 1 и 2). Обзор, однако, не является исчерпывающим, так как основан на опубликованной литературе по отдельным видам, личных наблюдениях авторов за видами и словесных описаниях М. Уорти [2] 4918 видов (около половины видов в Класс Авес). Этот автор основывал большинство своих описаний на обзоре литературы, который он перевел в шкалу темноты глаз с пятью категориями от желтого (0,00) до черного (1,00). Для большого числа видов, особенно менее изученных и/или обитающих в отдаленных районах, отсутствуют подробные сведения о цвете глаз особей обоих полов и/или разных возрастных классов. Глаза не сохранились у музейных экземпляров, и на этикетках обычно не упоминается окраска глаз во время поимки.
Из нашего, по общему признанию, ограниченного обзора мы определили, что, если цвет глаз различается в пределах одного вида, предсказуемое изменение с возрастом является наиболее частой причиной изменчивости, что согласуется с [16], и что этот способ изменчивости присутствует у всех видов. все существующие линии птиц (см. рис. 2), включая как неворобьиных (не менее 24 семейств), так и воробьиных (не менее 5 семейств). Цветовые переходы идут от темного оттенка к более светлому/яркому, от коричневого к желтому, от коричневого к красному или от желтого к красному, как описано в [17]. Направленность этих изменений может отражать последовательное вовлечение различных пигментов [4]: меланины будут более распространены у кареглазой молоди, в то время как другие пигменты, которые накапливаются с пищей, такие как каротиноиды [21], придают окраску кареглазым деткам. взрослый глаз. В большинстве случаев окраска глаз у диких животных остается неизменной. Особи определенного вида одного пола и возраста имеют одинаковую окраску глаз. Единственными животными, демонстрирующими изменчивость окраски глаз на взрослой стадии, являются домашние виды. Однако необходимы дальнейшие исследования, чтобы определить фактическую степень изменчивости (то есть оттенков и частот) у каждого домашнего вида.
Филогенетические отношения между 58 видами птиц с четырьмя категориями возрастных изменений цвета глаз (от коричневого к красному, от коричневого к желтому, от желтого к красному и от темного к светлому; изображено в подписи к вкладышу). Филогенетическое дерево представляет собой консенсусное дерево методом наименьших квадратов, рассчитанное на основе матрицы среднего святоотеческого расстояния набора из 1000 вероятных филогений из Jetz et al. 2012 (Природа 491: 444–448). Реконструкция наследственных состояний была сделана путем стохастического картирования признаков с использованием эмпирического подхода байесовских цепей Монте-Карло Маркова (MCMC) (Huelsenbeck et al. 2003, Systematic Biology 52: 131–158), реализованного в phytools (Revell 2012, Methods in Ecology and Evolution 3: 217–223)
Изображение в натуральную величину
Генетические детерминанты вариации цвета глаз
Окраска глаз человека наиболее разнообразна в Европе [22], но наблюдается наблюдаемая изменчивость внутри карих глаз в другом месте [23]. На самом деле цвет глаз человека постоянно варьируется от очень светло-голубого до очень темно-коричневого из-за различий в содержании меланина в радужной оболочке [24, 25].
На молекулярном уровне большая часть различий между светлой и темной окраской глаз у людей объясняется единственной мутацией в гене HERC2, которая влияет на выработку меланина в гене пигментации, известном как OCA2, и отвечает за карие глаза или не карие глаза [10, 23]. Тем не менее, недавние исследования показали, что другие гены влияют на пигментацию радужной оболочки. Например, первое полногеномное ассоциативное исследование с использованием количественных измерений цвета глаз по цифровым изображениям вместо категориальной вариации цвета глаз выявило три новых локуса, способствующих тонким различиям в естественном цвете глаз, подтверждая, что цвет глаз является полигенным признаком [25]. Известно не менее 50 SNP (однонуклеотидных полиморфизмов), связанных с изменением цвета глаз человека (https://www.snpedia. com/). Генетика изменчивости цвета глаз все еще находится в стадии интенсивного изучения, и уже очевидно, что цвет глаз у людей не является примером простого менделевского наследования, как утверждают старые учебники.
Молекулярная основа изменения окраски глаз у домашних животных указывает на то, что голубые глаза и другие фенотипы светлого цвета часто связаны с мутациями окраски шерсти, влияющими на выработку меланина. Например, ген розового глаза или локус p у мышей отвечает за светлые глаза и является ортологом гена P у людей, вызывающего альбинизм [26]. У диких животных генетическая информация скудна. Одно исследование [27] было направлено на то, чтобы определить, имеют ли голубые глаза диких лемуров ту же генетическую основу, что и голубые глаза человека, и было обнаружено, что последовательность гомологичного интрона гена HERC2 полностью сохранена у лемуров и других приматов (за исключением голубоглазые люди). Поэтому голубые глаза у человека и их дальних приматов-родственников лемуров — это фенотипическая конвергенция. Мы призываем к дальнейшим исследованиям того, как окраска глаз регулируется на генетическом уровне, чтобы понять давление отбора, действующее (или не действующее) на разные виды.
Последствия гипотезы
Как указано выше, только домашние животные имеют переменную окраску глаз. Светлые особи у темноокрашенных видов могут возникать как мутации «de novo», например, у коалы с голубыми глазами, которая недавно попала в заголовки газет как первая в истории Австралии (http://www.dailymail.co.uk/news/article). -508336/Meet-Frankie-rare-blue-eyed-baby-koala-named-Sinatra.html). Одомашненные животные значительно изменились по сравнению с исходным диким типом в окраске шерсти или оперения [28], а цвет глаз претерпел параллельный процесс. Изменение цвета глаз у домашней кошки, например, необычайно изменчиво: в некоторых диаграммах цвета глаз указано более 25 различных цветов глаз (например, http://aminoapps.com/page/warriors/1546176/epidemiology-eye-pigmentation). . Голубоглазые особи регулярно встречаются у всех домашних пород, от верблюдов и коз до лошадей (дополнительный файл 1: рисунок S1).
Половой диморфизм в окраске глаз относительно редко встречается у диких видов [5], и в нашем обзоре (Дополнительные файлы 1 и 2) мы сообщаем только о 25 видах в 8 семействах с сильным филогенетическим компонентом, так как было 9 видов уток. и 4 хищные птицы. Изменения окраски, связанные со статусом размножения, еще труднее обнаружить у диких позвоночных: нам известны только 5 видов птиц, включая бурого пеликана ( Pelecanus occidentalis ), у которого светлые глаза в период размножения совпадают с брачным периодом. оперение, но возвращается к более тусклому оперению и карим глазам в негнездовой период (https://identify.whatbird.com/obj/62/identification/Brown_Pelican.aspx). Уникальная ситуация среди млекопитающих связана с арктическим северным оленем ( Rangifer tarandus ), который сезонно меняет цвет глаз с золотистого на темно-синий из-за модификации Tapetum lucidum в ответ на зимнюю темноту [29]. Эти наблюдения приводят нас к постулату, что цвет радужной оболочки обычно не участвует в половом отборе у диких животных, а вместо этого может быть признаком естественного отбора. Однако, насколько нам известно, ни одно исследование не определило все факторы отбора, способствующие светлой или темной окраске глаз (но см. [5] для воробьиных).
Для домашних животных мы можем исключить половой отбор как движущую силу изменчивости, поскольку спаривание организуется и направляется владельцем-человеком, а естественный отбор также ослабляется благодаря защите от хищников и искусственному кормлению. Таким образом, мы предполагаем, что разнообразие цвета глаз у собак и других домашних животных напрямую связано с искусственным отбором. Предпочтение человеком редких фенотипов игнорирует даже сопутствующие негативные плейотропные эффекты [28]. Например, голубоглазые собаки или альпаки часто страдают глухотой, и им нелегко выжить в дикой природе.
Кажется, что у людей светлоокрашенные варианты возникли недавно, когда наш вид распространился в Европу из Африки [30]. Учитывая, что голубые глаза не дают явного преимущества с точки зрения возможностей зрения [31], существует вероятность того, что особи с цветом глаз, отличным от предположительно исходного фенотипа темных глаз, на самом деле предпочитались в качестве партнеров, таким образом быстро распространяя свои аллели. Это может быть случай полового отбора в действии, который, как известно, благоприятствует цветовым признакам и цветовым полиморфизмам [22].
Окраска глаз диких животных до сих пор остается малоизученной загадкой. Молекулярная основа окраски глаз человека изучена лучше, чем у любого другого таксона животных, но необходимы дальнейшие исследования, чтобы понять, обусловлена ли очевидная конвергентная эволюция множественной окраски глаз у людей и наших домашних видов одними и теми же или разными механизмами отбора.
Каталожные номера
Котт HB. Адаптивная окраска у животных. Лондон: Метуэн и Ко; 1940.
Google ученый
Уорти М. Цвета глаз животных: выбор авторов Press; 2000.
Бортолотти ГР. Естественный отбор и окраска: защита, сокрытие, реклама или обман? В: Hill GE, KJ MG, редакторы.
Окраска птиц, вып. 2: механизмы и измерения. Кембридж, Массачусетс: Издательство Гарвардского университета; 2006. с. 3–35.Google ученый
Олифант Л.В., Хадон Дж., Багнара Дж.Т. Рефугиумы пигментных клеток у гомойотермов. Уникальное эволюционное положение радужки. Пигментная клетка Res. 1992; 5: 367–71.
КАС Статья пабмед Google ученый
Дэвидсон Г., Тортон А., Клейтон Н.С. Эволюция цвета радужной оболочки в связи с гнездованием в полости и родительской заботой у воробьиных птиц. Биол Летт. 2017;13:20160783.
Артикул пабмед ПабМед Центральный Google ученый
Мэтисон И., Лазаридис И., Роланд Н., Маллик С., Паттерсон Н., Руденберг С.А., Харни Э., Стюардсон К., Фернандес Д., Новак, Бермудес де Кастро Дж.М., Карбонелл Э., Герритсен Ф.
, Хохлов А., Кузнецов П., Лозано М., Меллер Х., Мочалов О., Моисеев В., Рохо Герра М.А., Руденберг Дж., Мария Вержес Дж.М., Краузе Дж., Купер А., Альт К.В., Браун Д., Энтони Д., Лалуэза-Фокс С., Хаак В., Пинхаси Р., Райх D. Полногеномные закономерности отбора у 230 древних евразийцев. Природа. 2015;528:499-503.Антрогенные. Удивительная бледная пигментация в мезолитических HG Мотала, Форум. 2015 г. http://www.anthrogenica.com/showthread.php?3975-Surprising-Pale-pigmentation-in-Mesolithic-Motala-HGs. По состоянию на 24 ноября 2017 г.
Лазаридис И., Паттерсон Н., Миттник А., Рено Г., Маллик С. и др. Древние человеческие геномы предполагают наличие трех предковых популяций современных европейцев. Природа. 2013;513(7518):409–13.
Артикул Google ученый
Олальде И. и др. Получены иммунные и наследственные аллели пигментации у 7000-летнего мезолитического европейца.
Природа. 2014: 225–8.Eiberg H, Troelsen J, Nielsen M, et al. Голубой цвет глаз у людей может быть вызван идеально ассоциированной мутацией основателя в регуляторном элементе, расположенном в гене HERC2 , ингибирующем экспрессию OCA2 . Хам Жене. 2008; 123:177–87.
КАС Статья пабмед Google ученый
Higham T, et al. Время и пространственно-временные закономерности исчезновения неандертальцев. Природа. 2014;512(7514):306–9. 10.1038/природа13621.
КАС Статья пабмед Google ученый
Скоглунд П. и др. Древний геном волка обнаруживает раннее расхождение предков домашних собак и примесь высокоширотных пород. Карр Биол. 2015;25:1515–9.
КАС Статья пабмед Google ученый
Крейг AJFK, Hulley PE. Цвет радужной оболочки у воробьиных птиц: к чему быть яркоглазым? S Afr J Sci. 2004; 100: 584–8.
Google ученый
Бортолотти Г.Р., Смитс Дж., Бёрд Д.М. Цвет радужки у американских пустельг зависит от возраста, пола и воздействия ПХД. Физиол Биохим Зоол. 2003;76:99–104.
Артикул пабмед Google ученый
Уилсон Дж., Хартли И.Р. Изменения цвета глаз молодых бородатых синиц Panurus biarmicus и их использование для определения продуктивности размножения. Ибис. 2007; 149: 407–11.
Артикул Google ученый
Фрост П. Загадка европейского цвета волос, глаз и кожи. Ад Антропол. 2014; 4:78–88.
Артикул Google ученый
Frost P, Kleisner K, Flegr J. Состояние здоровья в зависимости от пола, цвета волос и цвета глаз: Рыжеволосые женщины наиболее разнообразны. bioRxiv. 2017.
Martinez-Cadenas C, Peña-Chilet M, Ibarrola-Villava M, Ribas G. Пол является основным фактором, объясняющим расхождения в предсказании цвета глаз на основе генотипа HERC2/OCA2 и модели IrisPlex. Судебно-медицинская экспертиза Int-Gen. 2013;7:453–60.
КАС Статья Google ученый
Фрост П. Европейский цвет волос и глаз. Случай частотно-зависимого полового отбора? Эвол Хам Бехав. 2006; 27:85–103.
Артикул Google ученый
Эдвардс М., Ча Д., Критика С., Джонсон М., Кук Г., Парра Э.Дж. Пигментация радужки как количественный признак: вариации в популяциях европейского, восточноазиатского и южноазиатского происхождения и связь с полиморфизмами генов-кандидатов. Пигментно-клеточная меланома Res. 2016;29:141–62.
КАС Статья пабмед Google ученый
Штурм Р.
А., Ларссон М. Генетика цвета и рисунка радужной оболочки человека. Пигментно-клеточная меланома Res. 2009 г.;22:544–62.КАС Статья пабмед Google ученый
Liu F, Wollstein A, Hysi PG, et al. Цифровая количественная оценка цвета человеческого глаза подчеркивает генетическую ассоциацию трех новых локусов. PLoSGenet. 2010;6(5):e1000934. 10.1371/журнал.pgen.1000934.
Google ученый
Brilliant M. Гены мыши p (конъюнктивальное разведение) и человека P, глазо-кожный альбинизм 2 типа (OCA2) и меланосомный рН. Пигментная клетка Res. 2001; 14:86–9.3.
КАС Статья пабмед Google ученый
Брэдли Б., Педерсен А., Манди Н.И. Голубые глаза у лемуров и человека: одинаковый фенотип, разные генетические механизмы. Am J Phys Антропол. 2009; 139: 269–73.
Артикул пабмед Google ученый
Reissmann M, Ludwig A. Плейотропные эффекты мутаций, связанных с окраской шерсти, у людей, мышей и других млекопитающих. Semin Cell Dev Biol. 2013; 24: 576–86.
КАС Статья пабмед Google ученый
Стоккан К.А., и др. Сдвигающиеся зеркала: адаптивные изменения ретинальных отражений к зимней темноте у арктических северных оленей. Proc R Soc B. 2013; 280:20132451.
Артикул пабмед ПабМед Центральный Google ученый
Темплтон, Арканзас. Из Африки снова и снова. Природа. 2002; 416:45–51.
КАС Статья пабмед Google ученый
Donnelly MP, et al. Общий вид области OCA2-HERC2 и пигментации. Хам Жене.
2012; 131:683–96.КАС Статья пабмед Google ученый
Амат Ф., Волленберг К.С., Венсес М. Корреляты цвета и рисунка глаз у мантелидных лягушек. Саламандра. 2013;49:7–17.
Google ученый
Nogueira DM, Alves MAS. Цвет радужной оболочки как показатель возрастного признака у самок бразильских танагров (Passeriformes: Emberizidae), подтвержденный методом молекулярного определения пола. Int J Trop Biol. 2008;56(4):629–1633.
Google ученый
Гальван И., Гарридо-Фернандес Х., Риос Х., Перес-Гальвес А., Родригес-Эррера Б., Негро Х.Х. Тропическая летучая мышь как модель метаболизма каротиноидов кожи у млекопитающих. ПНАС. 2016; 113:10932–7.
Артикул пабмед ПабМед Центральный Google ученый
Ссылки на скачивание
Благодарности
Мы благодарим CONACYT за творческий грант #454959 для MC Blázquez. IG и JJN признательны за проект CGL2015-67796-P от испанской компании MINECO. IG была поддержана стипендией Рамона и Кахала (RYC-2012-10237) от MINECO. Мы хотим поблагодарить за вклад двух анонимных рецензентов, которые значительно улучшили оригинальную рукопись как в концепции, так и в стиле.
Финансирование
Неприменимо.
Наличие данных и материалов
Неприменимо.
Информация об авторах
Авторы и организации
Estación Biológica de Doñana (CSIC), Avda. Americo Vespucio 26, 41092, Севилья, Испания
Juan J. Negro, M. Carmen Blázquez и Ismael Galván
Centro de Investigaciones Biológicas del Noroeste (CIBNOR), 23096, Ла-Пас, 903 M.900, B.C.0, B.
C.0 Бласкес
Авторы
- Хуан Дж. Негро
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- M. Carmen Blázquez
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ismael Galván
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
Вклады
Все три автора внесли равный вклад в эту работу. Все авторы прочитали и одобрили окончательный вариант рукописи.
Автор, ответственный за переписку
Переписка с Хуан Дж. Негро.
Декларация этики
Информация для авторов
JJN и IG исследуют происхождение и функцию окраски животных, уделяя особое внимание роли меланина и каротиноидных пигментов. MCB — эколог-эволюционист, исследующий особенности жизненного цикла рептилий засушливых зон.
Одобрение этики и согласие на участие
Неприменимо.
Согласие на публикацию
Не применимо.
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих интересов
Примечание издателя
Springer Nature сохраняет нейтралитет в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительные файлы
Дополнительный файл 1: Рисунок S1.
Изменение цвета глаз домашних коз (флоридская порода из Испании). На рисунке показаны две голубоглазые и две кареглазые особи из одного стада. (JPEG 3789kb)
Дополнительный файл 2: Таблица S1.
Исследование (неполное из-за нехватки опубликованной информации) изменений цвета глаз у диких видов птиц. Виды с продемонстрированными изменениями в окраске глаз составляют менее 1% всех существующих видов птиц, и в большинстве случаев изменения происходят, когда молодые птицы становятся взрослыми (см. , например, Nogueira DM, Alves MAS. Цвет радужки как индикатор возрастного признака). у самок бразильских танагров (Passeriformes: Emberizidae), подтвержденных методом молекулярного определения пола (Int. J. Trop. Biol. 2008; 56(4): 629).–1633). (XLSX 23 КБ)
Права и разрешения
Открытый доступ Эта статья распространяется в соответствии с условиями международной лицензии Creative Commons Attribution 4.0 (http://creativecommons.org/licenses/by/4.0/), которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии вы должным образом указываете автора (авторов) и источник, предоставляете ссылку на лицензию Creative Commons и указываете, были ли внесены изменения. Отказ от права Creative Commons на общественное достояние (http://creativecommons.org/publicdomain/zero/1.0/) применяется к данным, представленным в этой статье, если не указано иное.
Перепечатки и разрешения
Об этой статье
Выпученные глаза большехвостых граклов предлагают новый взгляд на птичье зрение
Новости
Это первый известный вид птиц, который смотрит на два объекта одновременно.
Энди Карстенс Репортер журнала Audubon
28 октября 2021 г.
Энди Карстенс Репортер, журнал Audubon
28 октября 2021 г.
Птицы в этой истории
Популярные истории
- Как отличить ворона от вороны
- Как приготовить нектар колибри
- Богомол против колибри
- Познакомьтесь с этими 20 распространенными птицами
- 13 забавных фактов о совах
Возможно, ни одна другая пара глаз в животном мире не так узнаваема, как у хамелеона. Мало того, что их большие выступающие глаза вращаются во всех направлениях, каждый глаз также может независимо осматривать свое окружение в поисках пищи и хищников, одновременно видя два отдельных объекта — талант, который, как считается, есть только у хамелеона.
Но, как оказалось, у большехвостых граклов тоже может быть этот навык. В исследовании, опубликованном прошлой весной в Experimental Brain Research г. ученые обнаружили, что граклы способны одновременно смотреть каждым глазом на разные изображения. До сих пор неизвестно, фокусировались ли граклы на двух изображениях и действительно ли регистрировали их одновременно — различие между «смотрением» и фактическим «видением» очень важно, — но результаты повышают такую вероятность, говорит Джессика Йорзински, специалист по поведению животных в Техасском университете A&M. Университет и автор исследования
Чтобы проверить, могут ли большехвостые граклы смотреть на разные объекты, Йорзински показывал два изображения человеческих лиц одновременно, причем каждое изображение отображалось на разных мониторах, расположенных вокруг птиц. Поскольку певчие птицы воспринимают людей как угрозу, лица вызывали сильную визуальную реакцию, которую Йорзински зафиксировал с помощью видеокамер.
Установив два монитора немного сзади, над или под птицами, Йорзински также смог максимизировать движения их глаз. Когда изображения вспыхивали, граклам приходилось отводить оба глаза от клюва и либо вверх, либо вниз, чтобы одновременно видеть человеческие лица. Проанализировав видео, она обнаружила, что птицы часто направляли одно глазное яблоко на каждую картинку одновременно.
Способность одновременно смотреть на два разных объекта зависит от того, как ориентированы глаза животного и могут ли они двигать ими независимо друг от друга. Глазные яблоки человека, как и у сов и большинства млекопитающих, расположены на передней части нашего лица, сцеплены вместе и сосредоточены на одной и той же области. Напротив, большехвостые граклы и большинство других птиц имеют боковые глаза, расположенные по бокам головы.
Пользы от такого навыка в дикой природе может быть много. «Один глаз может смотреть на что-то здесь, а другой глаз может смотреть на что-то там», — говорит Йорзински. «Это определенно потенциально очень полезная способность — иметь возможность двигать каждым глазом отдельно от другого». Это особенно верно, если птицы действительно могут отслеживать отдельные объекты. Если бы птица могла добывать пищу на земле одним глазом, а другим высматривая в небе хищников, это было бы эволюционной сверхспособностью, подобной людям, имеющим глаза на затылке.
Если бы птица могла добывать пищу на земле одним глазом, а другим высматривая в небе хищников, это было бы эволюционной сверхспособностью, сродни человеческим глазам на затылке.
Хотя важно указать разницу между тем, что значит смотреть на объект, и действительно регистрировать его, также стоит отметить, что некоторые птицы с латеральными глазами двигают ими в разных направлениях, не будучи по-настоящему независимыми. Например, 2009 г.Исследование, опубликованное в Journal of Experimental Biology , показало, что зрение Зебры Зяблика странным образом следует третьему закону движения Исаака Ньютона: на каждое действие существует равная и противоположная реакция. Когда один глаз поворачивается вверх и вперед, чтобы сфокусироваться на объекте, другой глаз автоматически перемещается вниз и назад, не фокусируясь ни на чем.
Неясно, регистрирует ли вращающийся назад глаз вьюрка то, что он видит, но равное и противоположное движение глаз может предотвратить образование пробелов на периферии птицы, сказал Ханс-Йоахим Бишоф, биолог на пенсии, автор исследования Zebra Finch. в электронном письме. Периферийные промежутки могут сделать их более уязвимыми для нападения хищников.
До сих пор не известно ни о каких других видах птиц, способных одновременно смотреть более чем на один объект, но некоторые другие виды, в том числе голуби и павлины, могут двигать глазами независимо друг от друга. Это указывает на то, что зрительные системы других птиц могут быть похожи на большехвостых граклов. «Не будет шоком узнать, что другие виды тоже способны на это», — говорит Йорзински. Необходимы дополнительные испытания, чтобы понять, насколько широко распространена эта черта у птиц.
«Они, безусловно, могут находиться в континууме», — говорит Йорзински, что может помочь объяснить различия между ее выводами и выводами Бишоф. Она говорит, что это также может варьироваться в зависимости от вида, когда они занимаются различными видами деятельности, такими как полет, поиск пищи или пение.
Что касается вопроса о том, действительно ли мозг большехвостого гракла может регистрировать две вещи одновременно, Йорзински говорит, что, возможно, некоторые птицы приспособились непрерывно сканировать свое окружение обоими глазами. Или они могут быстро переключать внимание между глазами, стробируя сознание туда и обратно.
Хотя вопросы остаются, результаты Йорзински предлагают новый взгляд на то, как видят птицы. Десятилетия назад исследователи думали, что птицы перемещают взгляд в основном за счет движения головы, и в значительной степени игнорировали то, как движение глаз влияет на зрение птиц. «Важно понять, как виды, отличные от нас самих, понимают окружающий их мир», — говорит Йорзински.