Кольчатые черви. Тип кольчатые черви фото
Кольчатые черви, или аннелиды – это беспозвоночные животные.
Считается, что они произошли от плоских червей и в отличие от них, имеют более совершенное строение.
Аннелиды дали начало развития моллюсков и членистоногих.
По образу жизни, можно выделить несколько групп аннелидов: Аннелиды, ведущие подземный образ жизни. Типичным представителем этого класса является дождевой червь.
кольчатые черви таблица фото
- Подземные аннелиды питаются перегнившими частичками растений, в земле роют норки, предпочитают теплую и влажную почву;
- Аннелиды, ведущие подводный образ жизни, свободно передвигаются по дну (червь нереида), так же делают норки в дне, (червь пескожил). Питаются морские аннелиды микроорганизмами, водорослями, ведут преимущественно хищнический образ жизни.
- Существуют черви кольчатого типа, ведущие паразитический образ жизни, например, пиявки.


Тело аннелидов состоит из головы, туловища, поделённого на сегменты и анальной лопасти. Каждый из видов имеет свои внешние особенности. Малощетинковые черви. У этих червей отсутствуют пародии на конечности. Головной отдел плохо выражен, у них нет глаз. Для размножения присутствует на теле специальный поясок.
строение кольчатых червей фото
На каждом сегменте тела расположено четыре пары щетинок. Такое строение идеально подходит для роющего образа жизни. Размеры малощетинковых колеблются от пяти миллиметров, до трех метров.
У этих аннелид присутствуют пародии на конечности, тело от нескольких миллиметров, до трех метров. Головной отдел хорошо развит, у некоторых видов присутствуют жабры. По бокам тела расположены частым рядом хитиновые щетинки.
многощетинковые черви фото
Данные аннелиды отличаются от других тем, что у них отсутствуют хитиновые щетинки и пародии на конечности, имеются две присоски.
пиявка фото
Для аннелидов характерно половое и бесполовое размножение:
- Половое размножение. Мужские и женские особи выбрасывают половые клетки, которые всплывают на поверхность воды, где происходит их слияние, в результате, образуются личинки, из них появляются новые особи. Таким образом, размножаются черви нереиды. Малощетинковые черви являются гермафродитами, для них свойственно внутреннее половое размножение. Яйца оплодотворяются в организме одной из особей, в специальном коконе, из которого выходят уже полноценные черви. Такое размножение свойственно для пиявок, дождевых червей.
- Бесполовое размножение. Размножение происходит следующим образом: тело червя делится на несколько частей, стенки сегментов лопаются, выделившиеся из них половые клетки попадают в воду, где происходит развитие личинки, а родительские особи погибают. Таким образом, размножаются кольчатые паразиты.
Аннелиды широко распространены по всему миру. Эти животные обитают на суше, в морях, озёрах, океанах, а некоторые могут поселиться на дереве, например, тропическая пиявка. Пиявки могут паразитировать в организме животных, попадая туда через озёрную воду.
Эти животные обитают на суше, в морях, озёрах, океанах, а некоторые могут поселиться на дереве, например, тропическая пиявка. Пиявки могут паразитировать в организме животных, попадая туда через озёрную воду.
- Аннелиды не переносят кислую среду обитания;
- Пиявки могут очень долгое время находиться без еды, примерно полтора года;
-
Среди аннелидов встречаются живородящие;
-
Малощетинковые – гермафродиты;
-
Аннелиды являются прародителями моллюсков;
-
Самый большой земельный червь обитает в Австралии, длинна его достигает до трех метров;
-
Ледяной червь погибает при температуре плюс пять градусов по Цельсию, а разлагается при комнатной температуре. Некоторые виды аннелид излучают свечение при размножении и защите.
Изучив данную тему, можно сказать, что черви кольчатого типа имеют огромное значение для экосистемы, являясь не только основной пищей для рыб и животных.
Дождевые черви имеют огромное значение для растений, так, как разрыхляя почву, происходит улучшенное поступление кислорода к их корням. Не обходится без аннелид и в медицине, пиявки незаменимы при лечении бесплодия, простатита, варикоза.
Тип — Кольчатые черви
Классы — Многощетинковые черви
Поясковые черви
Мизостомиды
Тип Кольчатые черви Урок 19. Внешнее строение и образ жизни кольчатых червей | Поурочные планы по биологии 6-7 класс
Цели урока: Рассмотреть особенности внешнего строения кольчатых
червей на примере дождевого червя — представителя малощетинковых червей и
нереиды — многощетинкового обитателя морского дна; продолжать формирование
умений вести наблюдения за животными, сравнивать их между собой для составления
общей характеристики типа кольчатых червей; умения работать с инструктивными
карточками, с учебником (пользоваться его рисунками в качестве источника
знаний).
Оборудование: таблица «Тип Кольчатые черви. Дождевой червь»; живые дождевые черви, пинцеты, лупы, кусочки репчатого лука, листы плотной бумаги; блюдечки, чашки Петри или куски стекла; инструктивные карты.
Ход урока
I. Организационный момент
II. Проверка усвоения материала темы «Круглые черви»
1. Карточки-задания для индивидуальной работы.
Чем различаются острица и аскарида?
Какие профилактические меры необходимы для предупреждения аскаридоза и энтеробиоза ?
Какой вред наносит больному аскарида человеческая? Какие экологические усло- вия необходимы для осуществления жизненного цикла аскариды человеческой?
С помощью цифр, данных в списке, укажите признаки, характерные для той или иной группы червей (А и Б): А. Плоские черви. Б. Круглые черви.
1.
Паразитический образ жизни. 6. Имеются присоски.
6. Имеются присоски.
2. Свободноживущие формы. 7. Имеется кожно-мускульный мешок.
3. Двусторонняя симметрия. 8. Полость тела заполнена жидкостью.
4. Тело состоит из члеников. 9. Гермафродиты.
5. Тело не имеет члеников. 10. Раздельнополые.
С помощью цифр, данных в списке, определите признаки, характерные для видов червей (А и Б). А. Бычий цепень. Б. Человеческая аскарида.
1. Тело сплющено в спинно- 5. Промежуточный хозяин — крупный
брюшном направлении. рогатый скот.
2.
Тело членистое и плоское.
3. Тело удлиненное, в поперечном 7. Имеются присоски с крючочками,
сечении имеет округлую форму. 8. Имеются ротовое и анальное отверстия.
4. Живет в кишечнике человека. 9. Имеется только ротовое отверстие.
2, Решение творческой задачи (класс работает в четверках, все группы могут одновременно, независимо друг от друга штурмовать одну задачу, можно устроить «конкурс идей» или группы работают над разными задачами).
1)
Существует
паразит, который обитает в мозговой ткани овец. У него сложный жизненный цикл,
и одна его стадия, именно половая, должна проходить в пищеварительном тракте
волка. Но где гарантия, что волк съест именно эту овцу — ведь пастухи зорко
следят за стадом, собаки охраняют его, да и сама овца не желает быть съеденной.
Подсказка 1 (на отдельных листочках дает учитель после предварительного обдумывания и обсуждения). В ходе эволюции закрепилось, что паразит использует ресурс поведения волков — волки едят овец.
Подсказка 2. Овца сама приходит к волку.
Ответ: Поскольку паразит живет в мозге, он в состоянии влиять на поведение овцы. Зараженные овцы теряют способность двигаться по прямой. Они идут только по дуге, вскоре отбиваются от стада и попадают волку на обед, а паразит — по месту назначения. Эта болезнь овец называется «овечья вертячка».
2) Есть паразит, который обитает в теле муравьев. Промежуточным его хозяином является корова. Каким образом паразит обеспечивает попадание «своего» муравья-хозяина в пищеварительную систему коровы?
Подсказка 1. Корова сама съедает муравья.
Подсказка
2. Ресурсы поведения: корова ест траву; муравьи ползают по траве.
Ответ:
паразит изменяет поведение муравья следующим образом: . муравей стремится
заползти на самый верх травинки. Корова захватывает его языком вместе с Зеленью.
Вместе с муравьем в пищеварительный тракт коровы попадает и паразит.
муравей стремится
заползти на самый верх травинки. Корова захватывает его языком вместе с Зеленью.
Вместе с муравьем в пищеварительный тракт коровы попадает и паразит.
3. Дайте общую характеристику типа Круглые черви. Назовите черты усложнения круглых червей по сравнению с плоскими червями. (1 ученик рассказывает перед классом).
III. Изучение нового материала
На теме «Круглые черви» наше знакомство с червями не заканчивается. Мы познакомились лишь с одной из двух условно выделенных групп. Представители плоских и круглых червей относятся к группе низших червей, а к высшим — относятся кольчатые черви, или кольчецы.
Итак, тип Кольчатые черви. В чем заключается их более высокая организация по сравнению с плоскими и круглыми червями?
/
О Выясните по рис. 79, чем кольчатые черви отличаются от плоских и круглых червей. Каких изображенных на рисунках 79, 80, 81 кольчатых червей вы видели в природе? Где живут эти черви?
Хорошо известным всем представителем кольчатых червей
является дождевой червь. Он обладает яркими отличительными признаками: тело
его очень сильно вытянуто; состоит из большого числа похожих друг на друга сегментов,
образованных кольцевидными перетяжками. Сегменты еще иначе называют кольцами,
отсюда и название кольчатые или кольчецы. Дождевой червь — обычный обитатель
почвы. Чем богаче почва органическими остатками, тем больше в ней червей.
Особенно много червей на поверхности земли бывает после дождя, т.к. вода
заливает воздухоносные поры в земле и т.д.
Он обладает яркими отличительными признаками: тело
его очень сильно вытянуто; состоит из большого числа похожих друг на друга сегментов,
образованных кольцевидными перетяжками. Сегменты еще иначе называют кольцами,
отсюда и название кольчатые или кольчецы. Дождевой червь — обычный обитатель
почвы. Чем богаче почва органическими остатками, тем больше в ней червей.
Особенно много червей на поверхности земли бывает после дождя, т.к. вода
заливает воздухоносные поры в земле и т.д.
Анализируя рисунки 80 и 81, можно прийти к следующим выводам: по положению мягкого тела трубочника, возможно, что это донный житель, который приспособлен к роющему образу жизни, а судя по красивым формам и яркой окраске серпулы и афродиты — это морские обитатели.
Познакомимся очень кратко с трубочником, серпулой и афродитой.
Заслушивание сообщений.
К этому типу относятся также нереида, пескожил,
томоптерис (изображения их показаны на таблице «Дождевой червь»). Нереида —
бродячее донное морское животное. В природе обитает несколько видов нереид. Эти
черви активно передвигаются по дну, схватывая пищу открытыми челюстями. Пескожил
живет в песочном грунте, где роет норки. Питается различными мелкими
организмами и полуразложившимися тканями растений и животных, постоянно
пропуская через кишечник песчаный грунт, содержащий питательные органические
остатки. При опасности быстро уходит в глубину норки. У то- моптериса, живущего
в толще воды, очень сильно увеличены боковые выросты члеников тела, а щетинки
превращены в широкие гребные лопасти. Тело томоптериса очень прозрачно, что
делает его менее заметным для врагов.
Нереида —
бродячее донное морское животное. В природе обитает несколько видов нереид. Эти
черви активно передвигаются по дну, схватывая пищу открытыми челюстями. Пескожил
живет в песочном грунте, где роет норки. Питается различными мелкими
организмами и полуразложившимися тканями растений и животных, постоянно
пропуская через кишечник песчаный грунт, содержащий питательные органические
остатки. При опасности быстро уходит в глубину норки. У то- моптериса, живущего
в толще воды, очень сильно увеличены боковые выросты члеников тела, а щетинки
превращены в широкие гребные лопасти. Тело томоптериса очень прозрачно, что
делает его менее заметным для врагов.
Известно около 8 тысяч видов кольчатых червей, обитающих в определенных средах.
— В каких средах обитают кольчецы? Какой образ жизни они ведут?
В настоящее время всех кольчатых червей делят на
несколько классов, среди которых широко распространены и известны три (см. схему «Классификация кольчатых червей»).
схему «Классификация кольчатых червей»).
Тип | Кольчатые черви (около 8000 видов) | ||
Класс | Малощетинковые или олигохеты (3100 видов) Почвенные и пресноводные жители | Многощетинковые или полихеты (5300 видов) Морские обитатели | Пиявки (300 видов) Пресноводные свободноживущие или паразиты |
Представители | дождевой червь, трубочник | нереида, афродита, пескожил, серпула | пиявки |
По сравнению с плоскими и круглыми червями кольчецы
стоят на более высокой ступени организации и имеют большое значение для
понимания эволюции высших беспозвоночных животных: моллюсков и членистоногих.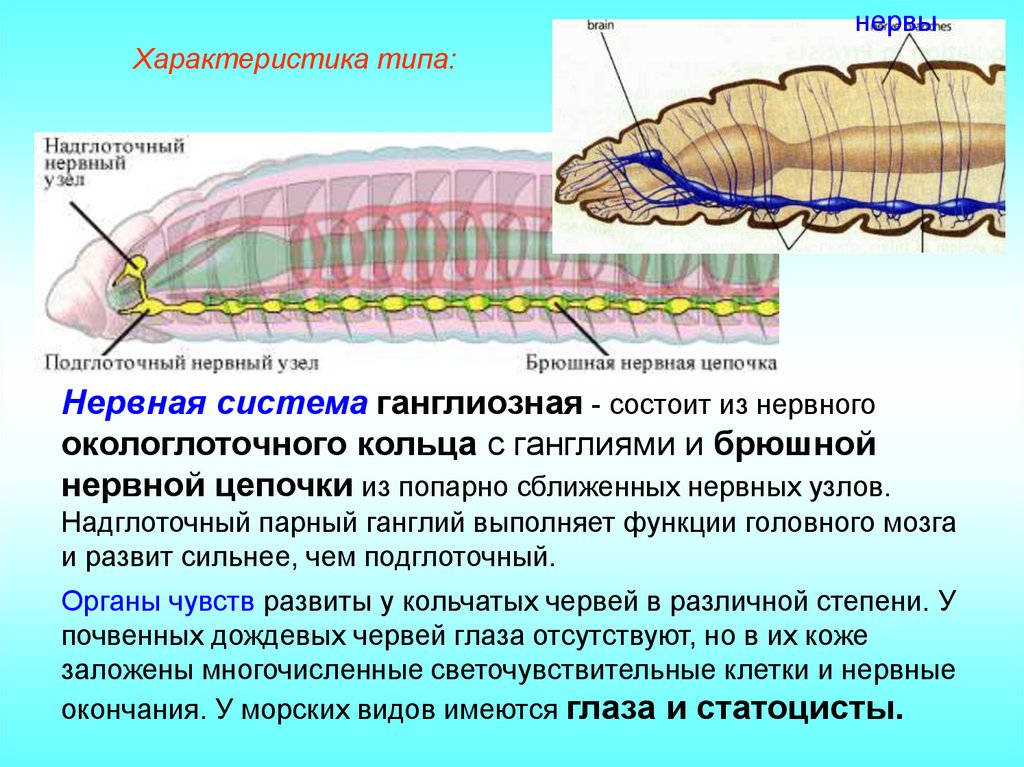 Чтобы дать общую характеристику червей этого типа, необходимо познакомиться с
особенностями их внешнего, внутреннего строения и процессов жизнедеятельности.
Чтобы дать общую характеристику червей этого типа, необходимо познакомиться с
особенностями их внешнего, внутреннего строения и процессов жизнедеятельности.
На примере дождевого червя рассмотрим строение малощетинковых червей. Попробуем добыть необходимые знания самостоятельно. А для этого выполним лабораторную работу «Внешнее строение дождевого червя, наблюдения за его передвижением и реакциями на раздражения», используем
живых дождевых червей. (Работа выполняется по инструктивной карточке, которую школьники получают вместе с животными и оборудованием).
Лабораторная работа №2 Внешнее строение дождевого червя, наблюдения за его передвижением и реакциями на раздражение
Инструктивная карточка
1. Рассмотрите живого дождевого червя, определите форму его тела, окраску, размеры (длину и толщину). Обратите внимание на его кольчатое (членистое) строение тела.
2.
По
движению червя определите передний и задний концы тела, их толщину и окраску.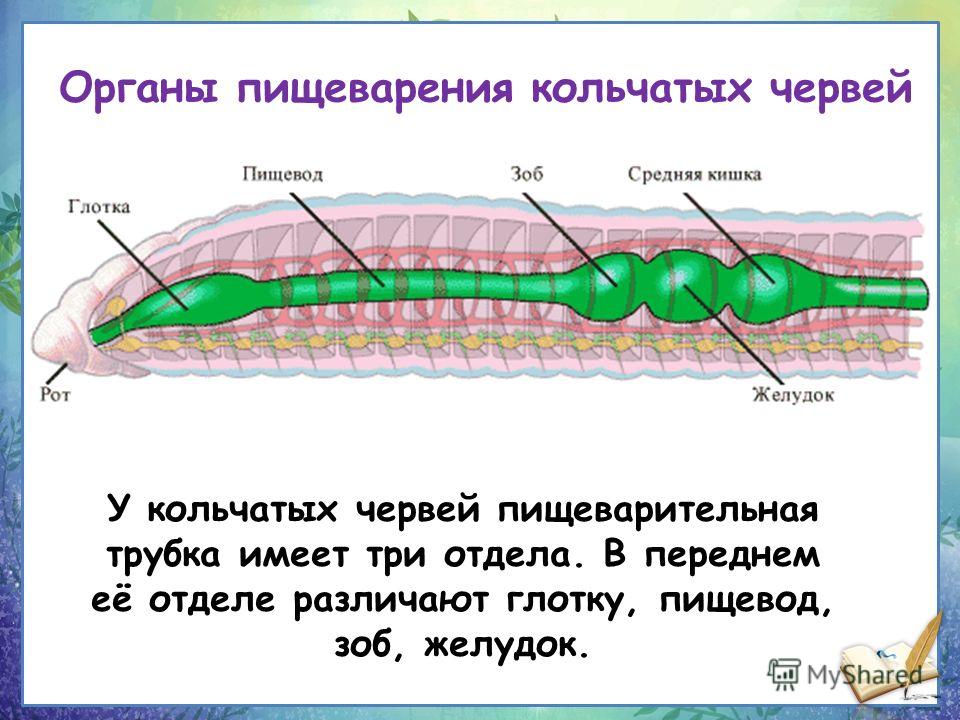
3. На переднем {более заостренном конце тела) найдите ротовое отверстие и утолщение — поясок. Подсчитайте, из скольких члеников он состоит.
4. На заднем (более тупом) конце тела найдите отверстие, через которое выбрасываются непереработанные остатки пищи, — это анальное отверстие.
5. Поверните червя брюшной стороной вверх. Определите, чем брюшная сторона червя отличается от спинной. Найдите с помощью лупы щетинки, которыми он упирается в частицы почвы при передвижении.
6. Пустите червя ползать по бумаге. Присмотритесь, как он передвигается. Прислушайтесь, как шуршит червь щетинками о бумагу. Смочите стекло водой и пустите червя на него. Посмотрите, как червь передвигается по мокрому стеклу.
7.
Проведите
пальцем вдоль брюшной стороны от переднего к заднему концу тела. Что ощущаете?
А теперь — от заднего к переднему и сравните степень сопротивляемости этим
движениям. Сделайте вывод.
Сделайте вывод.
8. Обратите внимание на кожу червя. Определите, какая она — сухая или влажная? Подумайте, какое значение имеет такая кожа для жизни в почве.
9. Выясните, как дождевой червь реагирует на действия различных раздражителей: прикоснитесь палочкой или пинцетом к различным участкам тела дождевого червя, поднесите к переднему концу тела кусочек лука, не дотрагиваясь до червя. Как реагирует червь на эти воздействия? Какие выводы можно сделать из проведенных наблюдений.
10. Опишите результаты наблюдений. Зарисуйте дождевого червя в тетради и сделайте надписи к рисунку. Ответьте на вопрос: каковы приспособления дождевого червя к условиям жизни в почве?
Результаты самостоятельной работы учащихся обсуждаются и формулируются выводы. Рисунки, подписи к ним и выводы можно оформить дома.
Черты приспособленности червя к обитанию в почве:
es 1. На каждом сегменте присутствует 4
пары щетинок, направленных назад, что позволяет червю цепляться всем телом за
неровности почвы и вместе с тем легко продвигаться вперед.
На каждом сегменте присутствует 4
пары щетинок, направленных назад, что позволяет червю цепляться всем телом за
неровности почвы и вместе с тем легко продвигаться вперед.
jss 2. Тело влажное, покрытое слизью, что защищает тело от грубых частиц почвы и облегчает передвижение.
jss 3. Органы чувств, расположенные на теле червя, помогают ориентироваться в земле, в окружающей среде.
О Используя рис. 82, сравним многощетинковых червей (нереида) с мало- щетинковыми (дождевой червь).
Таким образом, у многощетинковых червей впервые среди беспозвоночных животных наблюдается образование головного отдела (цефализация) и развитие на нем различных органов чувств. Также у них впервые возникают параподии (подвижные выросты стенки тела, усаженные щетинками) или примитивные конечности. Их рассматривают как зачаток более совершенной конечности. По видимому, из этих примитивных конечностей путем услож-
нения в процессе эволюции развились членистые
конечности членистоногих, («параподии» означает «похожие на ноги»).
IV. Закрепление
Какие выводы можно сделать из следующих фактов:
1. а) дождевые черви избегают сухой почвы и всегда держатся во влажной;
б) на поверхности почвы дождевые черви выползают из норок ночью либо днем после дождя?
2. На дождевого червя направили лупой солнечного «зайчика». Червь уходит от яркого света в темноту. К переднему концу тела поднесли смоченную в уксусе палочку. Червь отворачивается от палочки. Чем обусловлены реакции дождевого червя? Каково их биологическое значение?
V. Домашнее задание
§ 19, вопрос после §, термины; дооформить результаты лабораторной работы в тетради.
Влияние ископаемых данных на филогению кольчатых червей, выведенное из дискретных морфологических признаков
1. Rouse GW, Fauchald K.
1997.
Кладистика и полихеты. Зоол. Скр
26, 139–204. ( 10.1111/j.1463-6409.1997.tb00412.x) [CrossRef] [Google Scholar]
Скр
26, 139–204. ( 10.1111/j.1463-6409.1997.tb00412.x) [CrossRef] [Google Scholar]
2. Rouse G, Pleijel F. 2001. Полихеты. Оксфорд, Великобритания: Издательство Оксфордского университета. [Google Scholar]
3. Rouse GW, Pleijel F. 2007. Аннелида. зоотакса 1668, 245–264. [Google Scholar]
4. Rousset V, Pleijel F, Rouse GW, Erseus C, Siddall ME. 2007. Молекулярная филогения кольчатых червей. кладистика 23, 41–63. ( 10.1111/j.1096-0031.2006.00128.x) [CrossRef] [Google Scholar]
5. Zrzavý J, Riha P, Pialek L, Janouskovec J. 2009. Филогения кольчатых червей (Lophotrochozoa): общий анализ морфологии и шести генов. БМС Эвол. биол. 9, 189 (10.1186/1471-2148-9-189) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Struck T, Schult N, Kusen T, Hickman E, Bleidorn C, McHugh D , Галаныч К. 2007. Филогенез кольчатых червей и статус Sipuncula и Echiura. БМС Эвол. биол. 7, 57 ( 10.1186/1471-2148-7-57) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Макхью Д.
1997.
Молекулярные доказательства того, что эхиураны и погонофоры являются производными кольчатых червей. проц. Натл акад. науч. США
94, 8006–8009. ( 10.1073/pnas.94.15.8006) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Макхью Д.
1997.
Молекулярные доказательства того, что эхиураны и погонофоры являются производными кольчатых червей. проц. Натл акад. науч. США
94, 8006–8009. ( 10.1073/pnas.94.15.8006) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. McHugh D. 1999. Филогения кольчатых червей: Сиддалл и др. . (1998) опроверг. кладистика 15, 85–89. [Google Scholar]
9. Халаныч К.М., Дальгрен Т.Г., МакХью Д. 2002. Несегментированные кольчатые черви? Возможное происхождение четырех таксонов лофотрохозойских червей. интегр. Комп. биол. 42, 678–684. ( 10.1093/icb/42.3.678) [PubMed] [CrossRef] [Google Scholar]
10. Dordel J, Fisse F, Purschke G, Struck TH. 2010. Филогенетическое положение Sipuncula, полученное на основе мультигенных и филогеномных данных, и его значение для эволюции сегментации. Дж. Зул. Сист. Эвол. Рез. 48, 197–207. ( 10.1111/j.1439-0469.2010.00567.x) [CrossRef] [Google Scholar]
11. Struck T, et al.
2011.
Филогеномный анализ раскрывает эволюцию кольчатых червей.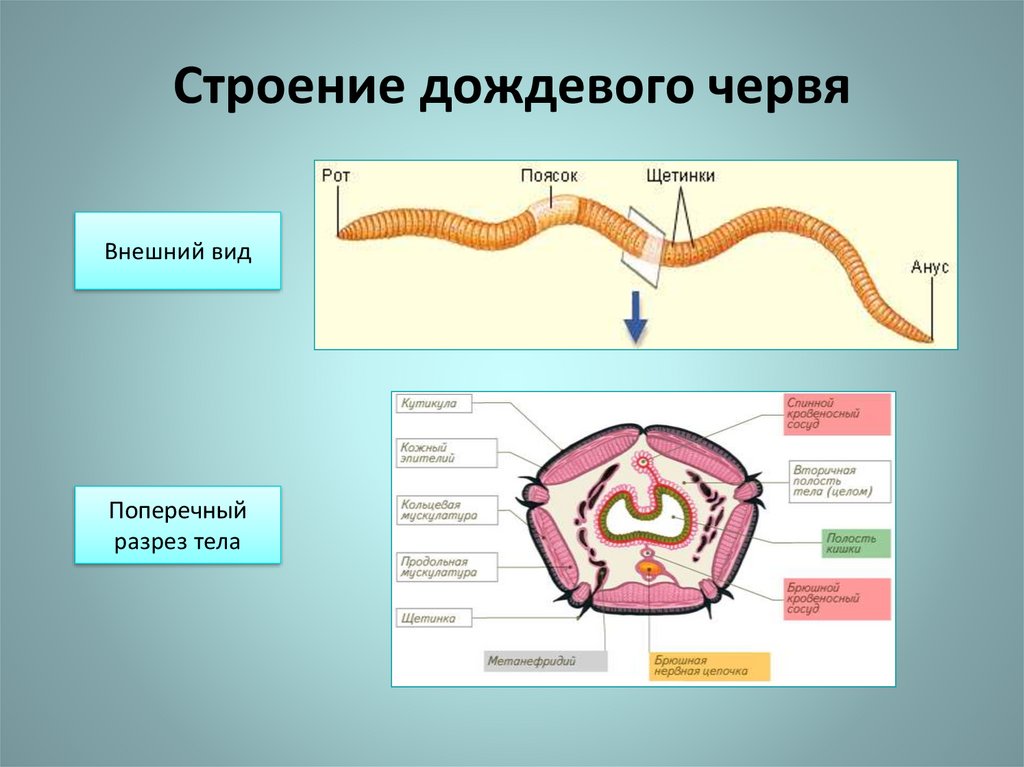 Природа
471, 95–113. ( 10.1038/nature09864) [PubMed] [CrossRef] [Google Scholar]
Природа
471, 95–113. ( 10.1038/nature09864) [PubMed] [CrossRef] [Google Scholar]
12. Weigert A, et al. 2014. Освещение основания дерева кольчатых червей с помощью транскриптомики. Мол. биол. Эвол. 31, 1391–1401. ( 10.1093/molbev/msu080) [PubMed] [CrossRef] [Google Scholar]
13. Андраде С.К., Ново М., Каваучи Г.Ю., Ворсаэ К., Плейел Ф., Гирибет Г., Роуз Г.В. 2015. Артикуляция «архианнелид»: филогеномика и отношения кольчатых червей с акцентом на таксоны мейофауны. Мол. биол. Эвол. 32, 2860–2875. ( 10.1093/molbev/msv157) [PubMed] [CrossRef] [Google Scholar]
14. Хачек Б. 1893 г. System der Anneliden, ein vorläufiger Bericht. Лотос 13, 123–126. [Google Scholar]
15. Hermans CO. 1969. Систематическое положение Archiannelida. Сист. биол. 18, 85–102. ( 10.1093/sysbio/18.1.85) [CrossRef] [Google Scholar]
16. Westheide W, McHugh D, Purschke G, Rouse G.
1999.
Систематизация кольчатых червей: разные подходы. В Репродуктивные стратегии и модели развития кольчатых червей. (редакторы AWC Dorresteijn, W Westheide), стр. 29.1–307. Берлин, Германия: Springer. [Google Scholar]
(редакторы AWC Dorresteijn, W Westheide), стр. 29.1–307. Берлин, Германия: Springer. [Google Scholar]
17. Вестхайде В. 1985. Систематическое положение Dinophilidae и проблема архианнелид. Происхождение и взаимоотношения низших беспозвоночных 397, 310–326. [Google Scholar]
18. Струк Т.Х., Голомбек А., Вейгерт А., Франке Ф.А., Вестхейде В., Пуршке Г., Блейдорн С., Халаныч К.М. 2015. Эволюция кольчатых червей выявляет два адаптивных пути к интерстициальной области. Курс. биол. 25, 1993–1999. ( 10.1016/j.cub.2015.06.007) [PubMed] [CrossRef] [Google Scholar]
19. Эйбай-Якобсен Д., Винтер Дж. 2012. Реконструкция предкового кольчатого червя. Дж. Зул. Сист. Эвол. Рез. 50, 85–87. ( 10.1111/j.1439-0469.2011.00651.x) [CrossRef] [Google Scholar]
20. Purschke G, Bleidorn C, Struck T. 2014. Систематика, эволюция и филогения кольчатых червей — морфологическая перспектива. Воспоминания Муз. Виктория 71, 247–269. [Google Scholar]
21. Вестхайде В.
1997.
Направление эволюции внутри многощетинковых червей. Дж. Нат. История
31, 1–15. ( 10.1080/00222939700770011) [CrossRef] [Google Scholar]
Дж. Нат. История
31, 1–15. ( 10.1080/00222939700770011) [CrossRef] [Google Scholar]
22. Кларк Р. 1964. Динамика эволюции многоклеточных животных : происхождение целома и сегментов. Оксфорд, Великобритания: Clarendon Press. [Google Scholar]
23. Баттерфилд, штат Нью-Джерси. 1990. Переоценка загадочной окаменелости сланца Берджесс Wiwaxia corrugata (Мэттью) и ее связи с полихетой Canadia spinosa Walcott. палеобиология 16, 287–303. ( 10.1017/S0094837300010009) [CrossRef] [Google Scholar]
24. Конвей Моррис С., Пил Дж. 2008. Самые ранние кольчатые черви: нижнекембрийские полихеты из Сириус Пассет Лагерстатте, Земля Пири, Северная Гренландия. Акта Палеонтол. Полоника 53, 135–146. [Google Scholar]
25. Liu J, Ou Q, Han J, Li J, Wu Y, Jiao G, He T. 2015. Нижнекембрийские полихеты из Китая проливают свет на раннюю эволюцию кольчатых червей. науч. Нац. 102, 1–7. ( 10.1007/s00114-014-1251-6) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Парри Л., Таннер А., Винтер Дж.
2014.
Происхождение кольчатых червей. Палеонтология
57, 1091–1103. ( 10.1111/pala.12129) [CrossRef] [Google Scholar]
Парри Л., Таннер А., Винтер Дж.
2014.
Происхождение кольчатых червей. Палеонтология
57, 1091–1103. ( 10.1111/pala.12129) [CrossRef] [Google Scholar]
27. Джонс Д., Томпсон И. 1977. Echiura из фауны пенсильванского Эссекса на севере Иллинойса. Летайя 10, 317–325. ( 10.1111/j.1502-3931.1977.tb00627.x) [CrossRef] [Google Scholar]
28. Bomfleur B, Kerp H, Taylor T, Moestrup O, Taylor E. 2012. Кокон триасовой пиявки из Антарктиды содержит ископаемое животное-колокольчик. проц. Натл акад. науч. США 109, 20 971–20 974. ( 10.1073/pnas.1218879109) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Манум С., Боуз М., Сойер Р. 1991. Ресничные коконы в пресноводных отложениях с триаса. Зоол. Скр. 20, 347–366. ( 10.1111/j.1463-6409.1991.tb00300.x) [CrossRef] [Google Scholar]
30. Edgecombe GD. 2010. Палеоморфология: окаменелости и вывод о кладистических отношениях. Акта Зоол. 91, 72–80. ( 10.1111/j.1463-6395.2009.00426.x) [CrossRef] [Google Scholar]
31. Донохью М.Дж., Дойл Дж.А., Готье Дж., Клюге А.Г., Роу Т.
1989.
Значение окаменелостей в реконструкции филогенеза. Анну. Преподобный Экол. Сист. 20, 431–460. ( 10.1146/annurev.es.20.110189.002243) [CrossRef] [Google Scholar]
Донохью М.Дж., Дойл Дж.А., Готье Дж., Клюге А.Г., Роу Т.
1989.
Значение окаменелостей в реконструкции филогенеза. Анну. Преподобный Экол. Сист. 20, 431–460. ( 10.1146/annurev.es.20.110189.002243) [CrossRef] [Google Scholar]
32. Sutton MD, Briggs DE, Siveter DJ, Siveter DJ. 2001. Трехмерно сохранившийся ископаемый многощетинковый червь из силурийского периода Херефордшира, Англия. проц. R Соц. Лонд. Б 268, 2355–2363. (10.1098/rspb.2001.1788) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Farrell U, Briggs D. 2007. Пиритизированная полихета из девона Онтарио. проц. Р. Соц. Б 274, 499–504. ( 10.1098/rspb.2006.0063) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Бриггс Д.Э., Бартелс К. 2010. Аннелиды из нижнедевонских хунсрюкских сланцев (нижний эмс, Рейнский массив, Германия). Палеонтология 53, 215–232. ( 10.1111/j.1475-4983.2009.00927.x) [CrossRef] [Google Scholar]
35. Vinther J, Eibye-Jacobsen D, Harper DA.
2011.
Раннекембрийская стеблевая полихета с пигидиальными усиками. биол. лат.
7, 929–932. ( 10.1098/rsbl.2011.0592) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
биол. лат.
7, 929–932. ( 10.1098/rsbl.2011.0592) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Eibye-Jacobsen D. 2004. Переоценка Wiwaxia и полихеты сланца Берджесс. Летайя 37, 317–335. ( 10.1080/00241160410002027) [CrossRef] [Google Scholar]
37. Parry L, Vinther J, Edgecombe GD. 2015. Кембрийские кольчатые черви стволовой группы и метамерное происхождение головы кольчатых червей. биол. лат. 11, 20150763 ( 10.1098 / rsbl.2015.0763) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Legg DA, Sutton MD, Edgecombe GD. 2013. Данные об окаменелостях членистоногих увеличивают соответствие морфологической и молекулярной филогении. Нац. коммун. 4, 2485 (10.1038/ncomms3485) [PubMed] [CrossRef] [Google Scholar]
39. Конуэй Моррис С., Пил Дж.С. 1995. Сочлененные халкирииды из нижнего кембрия Северной Гренландии и их роль в ранней эволюции первичноротых. Фил. Транс. Р. Соц. Лонд. Б 347, 305–358. ( 10.1098/rstb.1995.0029) [CrossRef] [Google Scholar]
40. Vinther J, Nielsen C.
2005.
Раннекембрийский Halkieria — моллюск. Зоол. Скр.
34, 81–89. ( 10.1111/j.1463-6409.2005.00177.x) [CrossRef] [Google Scholar]
Vinther J, Nielsen C.
2005.
Раннекембрийский Halkieria — моллюск. Зоол. Скр.
34, 81–89. ( 10.1111/j.1463-6409.2005.00177.x) [CrossRef] [Google Scholar]
41. Huang DY, Chen JY, Vannier J, Salinas JIS. 2004. Раннекембрийские сипункуловые черви из юго-западного Китая. проц. Р. Соц. Лонд. Б 271, 1671–1676. ( 10.1098/rspb.2004.2774) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Brazeau MD. 2011. Проблемные методы кодирования символов в морфологии и их последствия. биол. Дж. Линн. соц. 104, 489–498. ( 10.1111/j.1095-8312.2011.01755.x) [CrossRef] [Google Scholar]
43. Роуз Г. 2000. Полихеты много раз развивали питающихся личинок. Бык. мар. 67, 391–409. [Google Scholar]
44. Оррейдж Л., Мюллер М.К. 2005. Морфология нервной системы Polychaeta (Annelida). В морфологии, молекулах, эволюции и филогении полихет и родственных таксонов. (ред. Т. Бартоломеус, Г. Пуршке), стр. 79.–111. Берлин, Германия: Springer. [Google Scholar]
45. Пуршке Г.
2015.
24 Annelida: базальные группы и Pleistoannelida. В Строение и эволюция нервной системы беспозвоночных
(редакторы А. Шмидт-Реза, С. Харцш, Г. Пуршке), стр. 254–312. Оксфорд, Великобритания: Издательство Оксфордского университета. [Google Scholar]
Пуршке Г.
2015.
24 Annelida: базальные группы и Pleistoannelida. В Строение и эволюция нервной системы беспозвоночных
(редакторы А. Шмидт-Реза, С. Харцш, Г. Пуршке), стр. 254–312. Оксфорд, Великобритания: Издательство Оксфордского университета. [Google Scholar]
46. O’Reilly JE, Puttick MN, Parry L, Tanner AR, Tarver JE, Fleming J, Pisani D, Donoghue PC. 2016. Байесовские методы превосходят экономичность, но за счет точности оценки филогении по дискретным морфологическим данным. биол. лат. 12, 20160081 ( 10.1098/rsbl.2016.0081) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Райт А.М., Хиллис Д.М. 2014. Байесовский анализ с использованием простой модели правдоподобия превосходит экономию при оценке филогении по дискретным морфологическим данным. ПЛОС ОДИН 9, e109210 (10.1371/journal.pone.0109210) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Congreve CR, Lamsdell JC.
2016.
Предполагаемое взвешивание и его полезность в наборах палеонтологических данных: исследование с использованием смоделированных филогенетических матриц. Палеонтология
59, 447–462. ( 10.1111/pala.12236) [CrossRef] [Google Scholar]
Палеонтология
59, 447–462. ( 10.1111/pala.12236) [CrossRef] [Google Scholar]
49. Льюис PO. 2001. Вероятностный подход к оценке филогении по дискретным данным морфологических признаков. Сист. биол. 50, 913–925. ( 10.1080/106351501753462876) [PubMed] [CrossRef] [Google Scholar]
50. Goloboff PA, Farris JS, Nixon KC. 2008. TNT, бесплатная программа для филогенетического анализа. кладистика 24, 774–786. ( 10.1111/j.1096-0031.2008.00217.x) [CrossRef] [Google Scholar]
51. Ronquist F, et al. 2012. MrBayes 3.2: эффективный байесовский филогенетический вывод и выбор модели в большом модельном пространстве. Сист. биол. 61, 539–542. ( 10.1093/sysbio/sys029) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Стаматакис А. 2014. RAxML версии 8: инструмент для филогенетического анализа и пост-анализа больших филогений. Биоинформатика 30, 1312–1313. ( 10.1093/bioinformatics/btu033) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Hammer Ø, Harper D, Ryan P.
2001.
ПРОШЛОЕ — ПАлеонтологическая статистика, вер. 1,89. Палеонтол. Электрон.
4, 1–9. [Google Scholar]
Hammer Ø, Harper D, Ryan P.
2001.
ПРОШЛОЕ — ПАлеонтологическая статистика, вер. 1,89. Палеонтол. Электрон.
4, 1–9. [Google Scholar]
54. Уорсаэ К., Нигрен А., Роуз Г.В., Гирибет Г., Перссон Дж., Сандберг П., Плейел Ф. 2005. Филогенетическое положение Nerillidae и Aberranta (Polychaeta, Annelida), проанализированное путем прямой оптимизации объединенных молекулярных и морфологических данных. Зоол. Скр. 34, 313–328. ( 10.1111/j.1463-6409.2005.00190.x) [CrossRef] [Google Scholar]
55. Ваннингер А., Куп Д., Бромхэм Л., Нунан Э., Дегнан Б.М. 2005. Развитие нервной и мышечной системы у Phascolion strombus (Sipuncula). Дев. Гены Эвол. 215, 509–518. ( 10.1007/s00427-005-0012-0) [PubMed] [CrossRef] [Google Scholar]
56. Цетлин А., Пуршке Г. 2006. Тонкое строение глоточного аппарата личинки пелагосферы Phascolosoma agassizii (Sipuncula) и его филогенетическое значение. Зооморфология 125, 109–117. ( 10.1007/s00435-006-0025-x) [CrossRef] [Google Scholar]
57. Sansom RS, Gabbott SE, Purnell MA.
2010.
Неслучайный распад признаков хордовых вызывает предвзятость в интерпретации окаменелостей. Природа
463, 797–800. ( 10.1038/nature08745) [PubMed] [CrossRef] [Google Scholar]
Sansom RS, Gabbott SE, Purnell MA.
2010.
Неслучайный распад признаков хордовых вызывает предвзятость в интерпретации окаменелостей. Природа
463, 797–800. ( 10.1038/nature08745) [PubMed] [CrossRef] [Google Scholar]
58. Фицхью К., Срока С., Крути М., Хендерсон А., Хэй А. 1997. Многощетинковые черви. В справочнике Ричардсона по ископаемой фауне Мейзон-Крик. (редакторы CW Shabica, AA Hay), стр. 64–83. ДеКалб, Иллинойс: Издательство Северо-восточного университета Иллинойса. [Академия Google]
59. Винс Дж.Дж. 2005. Могут ли неполные таксоны спасти филогенетический анализ от притяжения длинных ветвей? Сист. биол. 54, 731–742. ( 10.1080/10635150500234583) [PubMed] [CrossRef] [Google Scholar]
60. Мерц Р.А., Вудин С.А. 2006. Многощетинковые щетинки: функции, окаменелости и филогения. интегр. Комп. биол. 46, 481–496. ( 10.1093/icb/icj057) [PubMed] [CrossRef] [Google Scholar]
Июнь — Обнаружен самый старый родственник мотыльковых и дождевых червей
Ученые обнаружили самое древнее ископаемое, которое можно отнести к ныне живущим кольчатым червям, группе животных который содержит дождевых червей, пиявок и многих других форм в океане, включая многощетинковых червей (таких как тряпичные черви и улитки).
Этот вид был обнаружен в месте, богатом раннекембрийскими окаменелостями, датируемыми примерно 514 миллионами лет назад, в восточной провинции Юньнань, Китай.
Эти исключительно хорошо сохранившиеся кембрийские породы являются важными ископаемыми свидетельствами зарождения ранней жизни животных, известного как «кембрийский взрыв».
Однако, несмотря на то, что кольчатые черви (сегментарные черви) являются одной из основных групп животных, ныне живущих, их окаменелости чрезвычайно редки среди этих самых ранних сообществ животных, поэтому ведущие ученые не уверены в их происхождении и ранней эволюции.
В статье, опубликованной сегодня в журнале Nature , международная группа ученых из Эксетерского, Юньнаньского, Оксфордского и Бристольского университетов описывает новый вид под названием Dannychaeta tucolus.
Они показывают, что он принадлежит к живой группе многощетинковых червей, называемых Maglonidae или лопатовидными червями.
В отличие от других кембрийских видов многощетинковых червей, Dannychaeta tucolus вел сидячий (фиксированный на одном месте) образ жизни внутри трубки, образ жизни, который у кольчатых червей известен с уверенностью только много миллионов лет спустя.
Многие группы современных кольчатых червей ведут оседлый образ жизни либо в защитных трубах, либо прячась от хищников в норах.
Биологи, работающие с живыми видами, утверждают, что эти сидячие кольчатые черви представляют собой очень древние ветви древа жизни кольчатых червей.
Однако оседлый образ жизни еще не был обнаружен в ранних окаменелостях кольчатых червей.
Maglonidae (живые лопатообразные черви) обитают в океанах по всему миру, в том числе в прибрежных районах Соединенного Королевства.
Соавтор, доктор Люк Пэрри из Оксфордского университета, сказал: «Живые кольчатые черви делают в современном океане самые разные вещи, например живут как сидячие фильтраторы и хищники из засады. Все древние кольчатые черви, о которых мы знали ранее в кембрии, скорее всего, ползали по морскому дну, и то, что мы видим в Dannychaeta , весьма радикально отличается.
Все древние кольчатые черви, о которых мы знали ранее в кембрии, скорее всего, ползали по морскому дну, и то, что мы видим в Dannychaeta , весьма радикально отличается.
«Обнаружение Dannychaeta говорит нам о том, что даже в очень ранних экосистемах, где доминировали животные, древние кольчатые черви выполняли многие из тех же функций, что и сегодня в океане».
Аспирант Хонг Чен, соавтор из Юньнаньского университета, сказал: «Мы были очень удивлены, обнаружив многощетинкового червя 514 миллионов лет назад, который жил в трубке, тем более что он так похож на виды, которые все еще живы. Cегодня.»
Ответственный автор, доктор Сяоя Ма из Центра экологии и охраны природы Эксетерского университета, сказал: «Это самое раннее ископаемое свидетельство сидячих кольчатых червей, а также первое появление живой группы кольчатых червей. Учитывая, насколько редки окаменелости кольчатых червей в раннекембрийский период, мы удивлены и восхищены этим открытием.