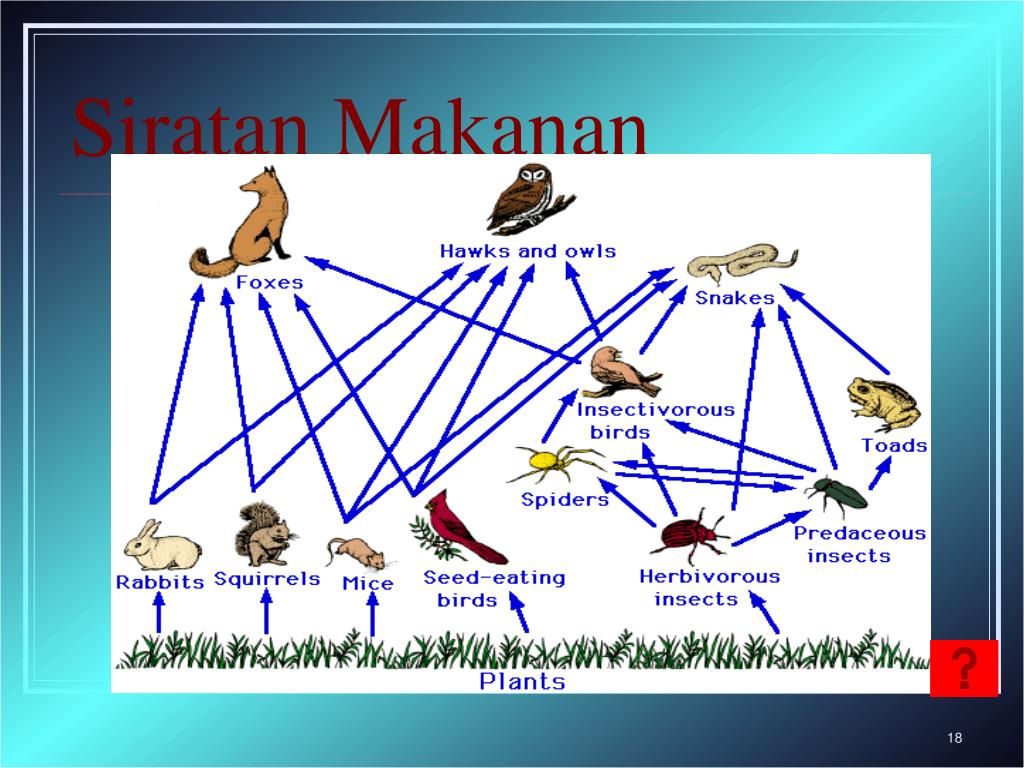
Пищевая цепочка: что это такое, характеристики и значение
Во всех естественных экосистемах существует экологический баланс и поток энергии между физической средой и живыми существами. Он известен как пищевая цепочка, пищевая цепь или пищевая цепь к этому механизму передачи органического вещества и энергии через различные виды живых существ, составляющих биологическое сообщество или экосистему. Это делает пищевую цепочку одним из важнейших элементов экосистем для их выживания и развития.
Поэтому мы собираемся посвятить эту статью, чтобы рассказать вам обо всех характеристиках, важности и типах пищевой цепи.
Индекс
- 1 Características principales
- 2 Пищевая цепочка
- 3 Типы пищевой цепи
Características principales
Необходимо учитывать, что все биологические сообщества состоят из разных форм жизни. Различные живые существа, населяющие экосистему, связаны друг с другом и со своей физической средой.
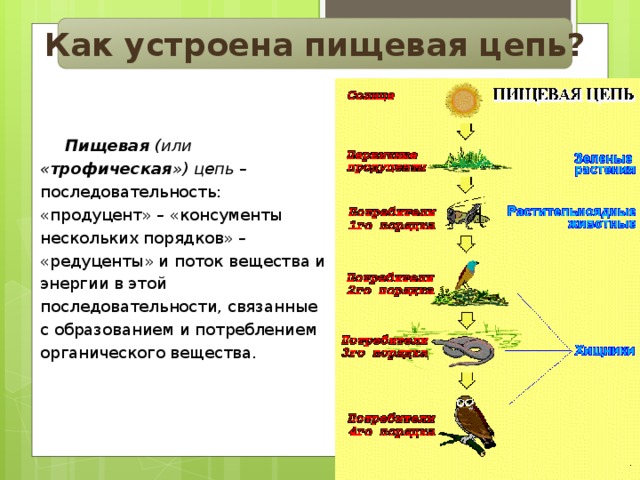
Пищевая цепочка состоит из различных звеньев в зависимости от типа функции, которую выполняет каждое живое существо. Таким образом, мы можем говорить о производителях, потребителях и переработчиках в пищевой цепи. Давайте посмотрим, какие различные функции выполняют живые существа в пищевой цепи:
- производители: они отвечают за питание остальных живых существ, используя неорганическую материю и источник энергии, такой как солнечный свет, для собственного развития.
 Например, посредством фотосинтеза растения могут поглощать питательные вещества. Растения служат пищей для других живых существ.
Например, посредством фотосинтеза растения могут поглощать питательные вещества. Растения служат пищей для других живых существ. - Потребительское право : они питаются органическим веществом других живых существ. Эти живые существа могут быть производителями или другими потребителями. В этом случае они будут действовать как хищники. В зависимости от случая мы можем называть потребителей первичными и вторичными.
- Разложители: Это те, кто отвечает за потребление разлагающегося органического вещества, чтобы восстановить его до самых элементарных компонентов. Эти деструкторы — это в основном грибы, насекомые-бактерии, присутствующие в экосистемах.
 Например, посредством фотосинтеза растения могут поглощать питательные вещества. Растения служат пищей для других живых существ.
Например, посредством фотосинтеза растения могут поглощать питательные вещества. Растения служат пищей для других живых существ.Пищевая цепочка
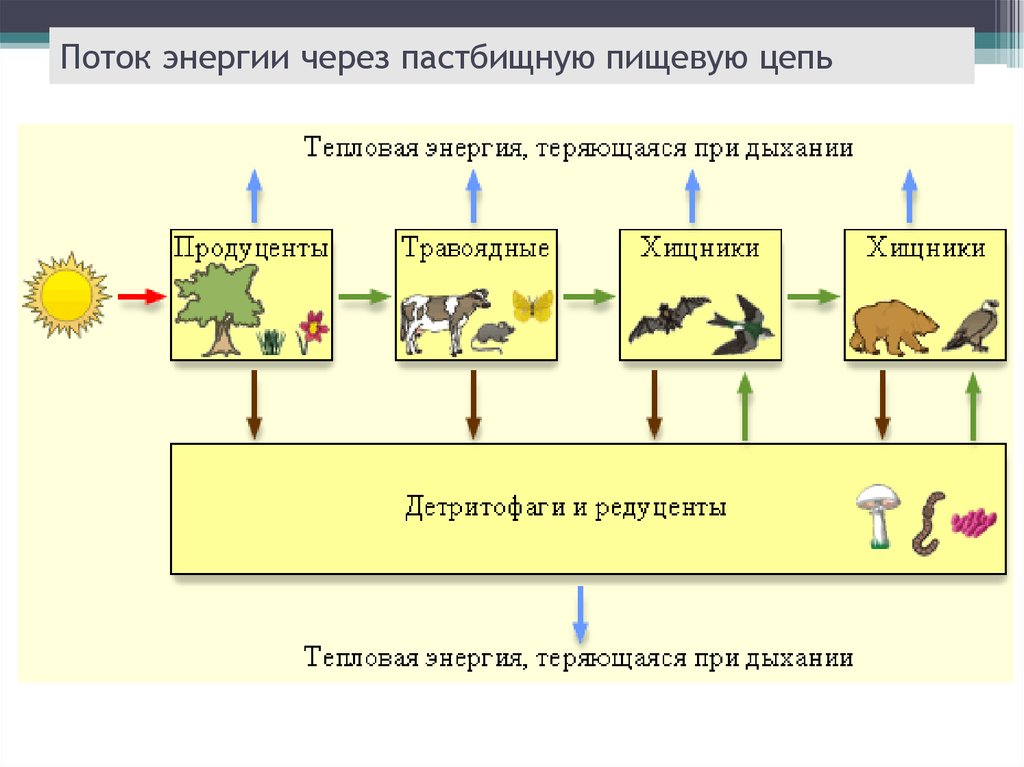
При этом необходимо учитывать, что для того, чтобы добраться до конечного потребителя, в переносе вещества приходится часть тепла. Каждое звено пищевой цепи или уровней — это цикл, поддерживающий определенный баланс. Проблема пищевой цепи — это человек. Вмешательство человека или какое-либо стихийное бедствие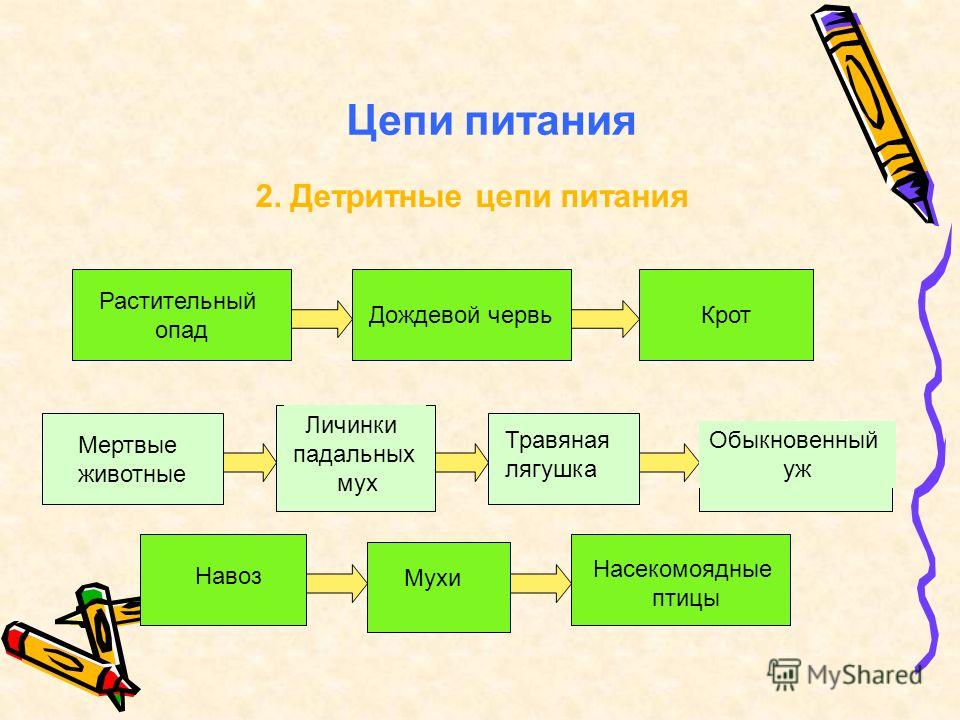 Они также способны нанести долгосрочный экологический ущерб.
Они также способны нанести долгосрочный экологический ущерб.
Наиболее естественные воздействия на пищевую цепь экосистемы происходят из-за инвазивных видов. Эти виды вытесняют местные виды. Это также может произойти, когда один из основных хищников уничтожен, чтобы предотвратить беспорядочное размножение других более мелких видов. Что касается людей, сокращение популяции иберийского волка вызывает рост кроликов. Именно эти кролики наносят ущерб урожаю.
Существуют пищевые цепи, в которых определенный процент энергии теряется при переходе от одного звена к другому. Это самый распространенный. Следовательно, когда достигается последнее звено потребителя, значительная часть передачи материи теряется. В свою очередь, химическая энергия передается от одной ткани к другой. Волк не ест траву, но он ест кроликов, которые, в свою очередь, едят траву. Энергия травы достигла волка в преобразованном виде. Хотя волк по пути потерял часть своей энергии, в конце концов волк поглощает его свойства.
Типы пищевой цепи
Мы собираемся указать, какие основные типы пищевых цепочек существуют. По среде обитания, в которой они встречаются, можно говорить о двух типах трофических цепочек:
- Наземные пищевые цепи: Это те, что имеют место в районах континентального шельфа. Они также могут возникать под поверхностью земли. Например, мы находим трофические кальдеры пустыни, тропического леса и т. Д.
- Водные пищевые цепи: те, которые происходят в морской или озерной среде. Обычно они состоят из существ, приспособившихся к водной жизни. Также можно найти подводные районы, где развивается трофическая цепочка. К водным также относятся прибрежные зоны и абиссальные зоны.
Трофические уровни — это уровни, которые указывают на различные части производства и обмена вещества. Каждая ступень пищевой цепи называется трофическим уровнем. Различные виды, отвечающие за совместное использование питательной активности и режима питания, — это те виды, которые занимают весь пищевой цикл экосистемы.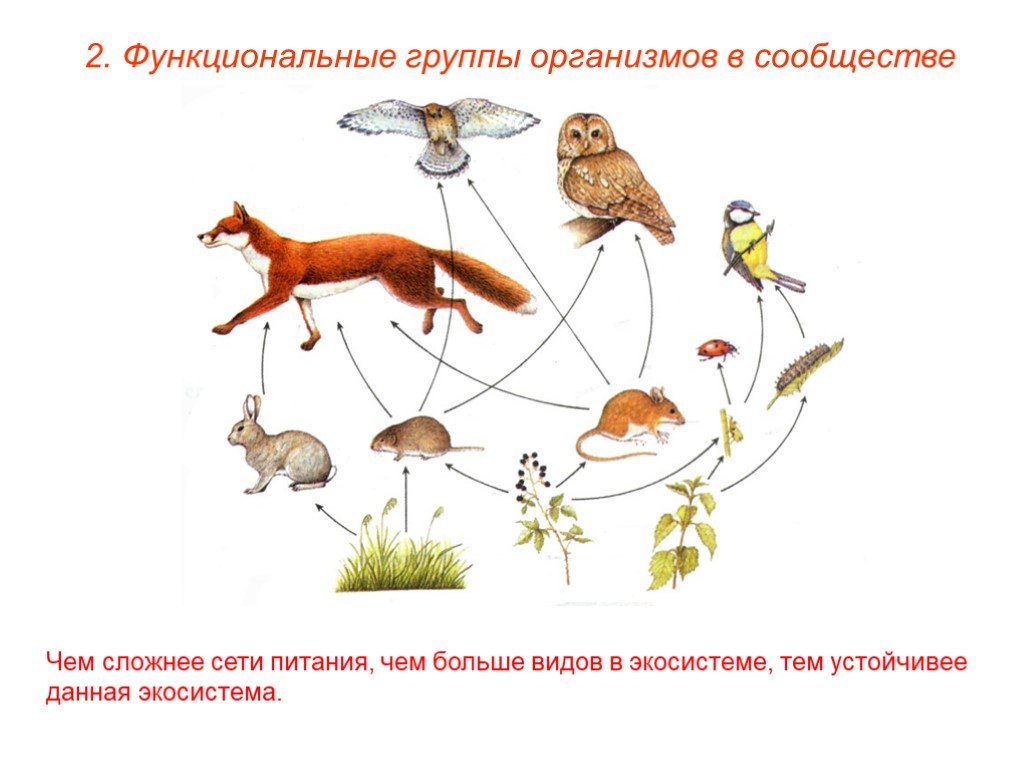
Это означает, что различные трофические уровни могут быть следующими:
- производители: это те формы жизни, которые можно воспитать автотрофным способом. Это означает, что они способны синтезировать собственную пищу.
- Потребители: это те живые и гетеротрофные существа, которые должны потреблять органическое вещество, чтобы питаться. Они делятся на следующие 4 ступени: первичную, вторичную, третичную и четвертичную. Первичные из них — те травоядные, которые питаются продуцентами. Вторичные — это те, которые питаются другими основными потребителями, но являются мелкими хищниками. Третичные хищники — более крупные хищники, которые охотятся на вторичных потребителей. Или четвертичные — это крупные хищники, которые питаются третичными или вторичными потребителями и не имеют естественных хищников.
- Разложители: можно сказать, что это природный отдел утилизации. Питается отходами и разлагающимися органическими веществами.
Я надеюсь, что с этой информацией вы сможете больше узнать о пищевой цепи и ее важности для экосистем.
детритная, пастбищная и паразитическая цепи питания окружающего мира
Биология
12.11.21
15 мин.
Пищевой, или трофической цепью называют взаимоотношение между различными группами организмов (растениями, грибами, животными и микробами), в котором происходит транспорт энергии в результате употребления в пищу одних особей другими. Перенос энергии — основа нормального функционирования экосистемы. Наверняка эти понятия знакомы вам с 9 класса школы из курса общей биологии.
Оглавление:
- Виды пищевых взаимоотношений
- Трофические уровни
- Значение пищевых взаимосвязей
Пищевой, или трофической цепью называют взаимоотношение между различными группами организмов (растениями, грибами, животными и микробами), в котором происходит транспорт энергии в результате употребления в пищу одних особей другими.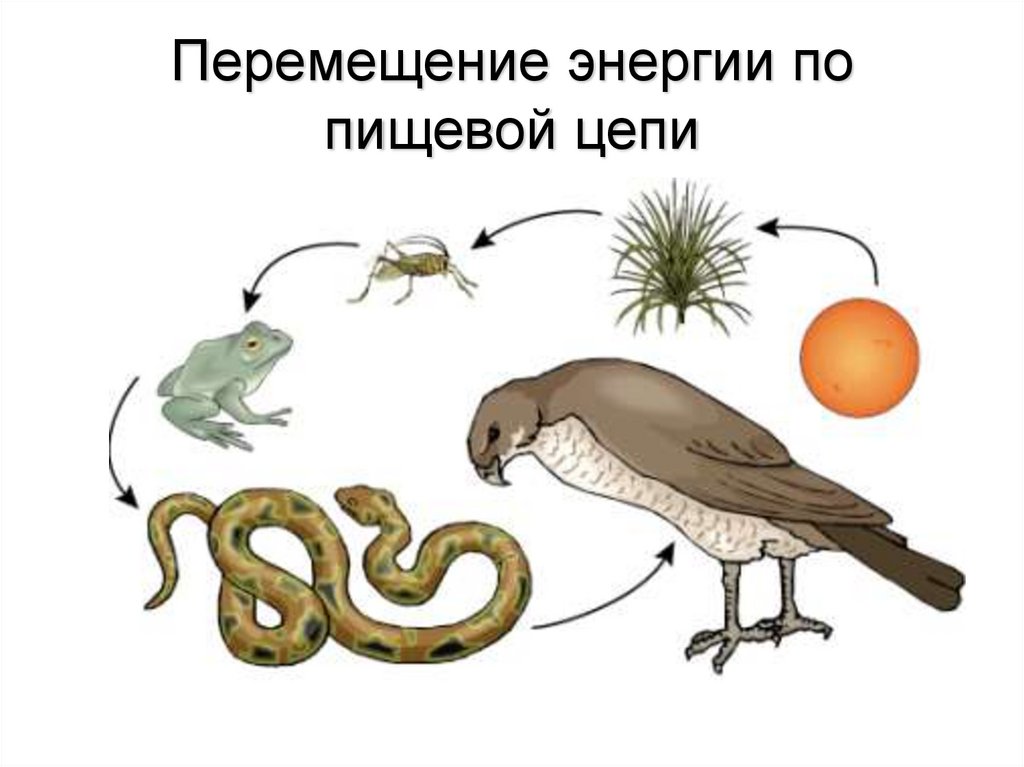 Перенос энергии — основа нормального функционирования экосистемы. Наверняка эти понятия знакомы вам с 9 класса школы из курса общей биологии.
Перенос энергии — основа нормального функционирования экосистемы. Наверняка эти понятия знакомы вам с 9 класса школы из курса общей биологии.
Особи последующего звена съедают организмы прошлого звена, и так происходит транспорт вещества и энергии по цепочке. Эта последовательность процессов лежит в основе живого круговорота веществ в природе. Стоит сказать, что огромная часть потенциальной энергии (примерно 85%) теряется при переносе от одного звена к другому, она диссипируется, то есть рассеивается в виде тепла. Этот фактор является лимитирующим по отношению к длине пищевых цепей, которые в природе обычно насчитывают 4—5 звеньев.
Виды пищевых взаимоотношений
Внутри экосистем органические вещества производятся автотрофами (продуцентами). Растения, в свою очередь, поедаются растительноядными животными (консументами первого порядка), которых затем съедают хищные животные (консументами второго порядка).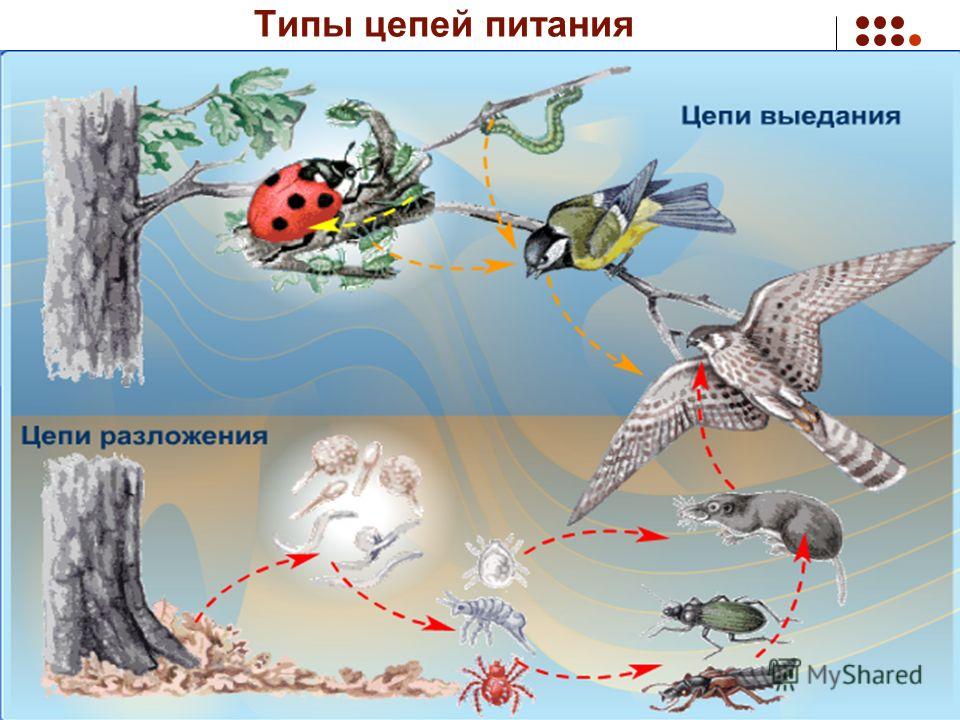
Различают:
- Пастбищные пищевые цепочки;
- Детритные пищевые цепочки;
- Паразитические пищевые цепочки.
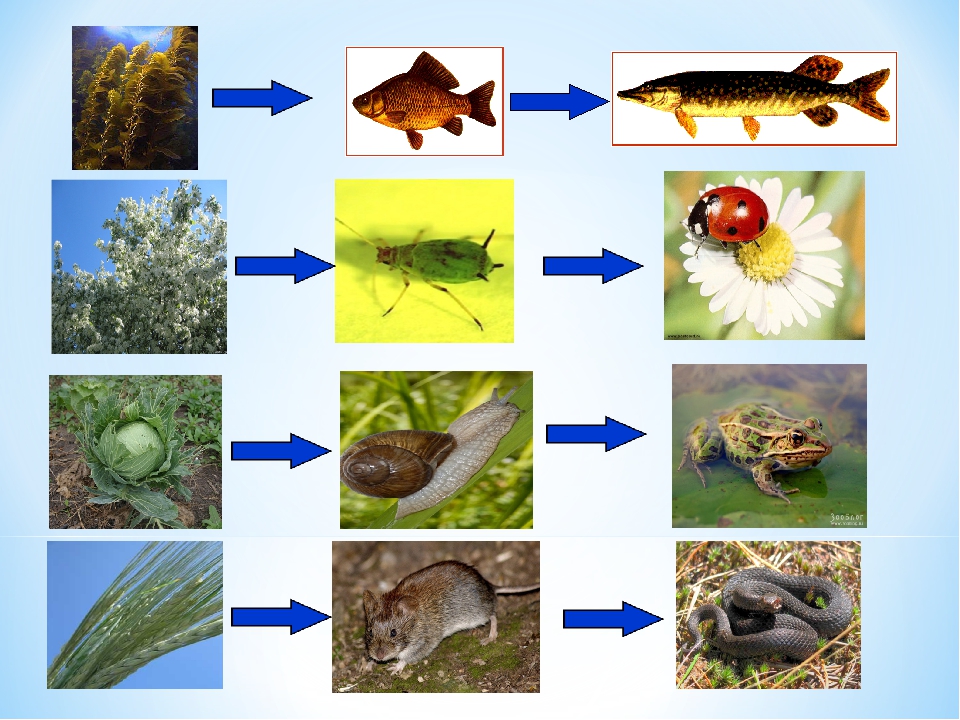
Пастбищные цепи
Трофические цепи начинаются с авто- или хемотрофов (продуцентов) и включают гетеротрофов в виде консументов различных порядков. Такие пищевые цепи широко распространены в сухопутных и морских экосистемах. Их можно нарисовать и составить в виде схемы:
Продуценты —> Консументы I порядка —> Консументы I. I. порядка—> Консументы III порядка.
Типичным примером является пищевая цепь луга (это может быть и лесная зона, и пустыня, в этом случае будут отличаться лишь биологические виды различных участников трофической цепи и разветвлённость сети пищевых взаимодействий).
Итак, цветок с помощью энергии Солнца производит для себя питательные вещества, то есть является продуцентом и первым звеном в цепи. Бабочка, которая питается нектаром этого цветка — консумент I порядка и второе звено. Лягушка, также обитающая на лугу и являющаяся насекомоядным животным, съедает бабочку — третье звено в цепи, консумент II порядка. Лягушку проглатывает уж — четвёртое звено и консумент III порядка, ужа съедает ястреб — консумент IV порядка и пятое, как правило, последнее звено в пищевой цепочке. Человек может присутствовать в этой цепи также в роли консумента.
Бабочка, которая питается нектаром этого цветка — консумент I порядка и второе звено. Лягушка, также обитающая на лугу и являющаяся насекомоядным животным, съедает бабочку — третье звено в цепи, консумент II порядка. Лягушку проглатывает уж — четвёртое звено и консумент III порядка, ужа съедает ястреб — консумент IV порядка и пятое, как правило, последнее звено в пищевой цепочке. Человек может присутствовать в этой цепи также в роли консумента.
В водах Мирового океана автотрофы, представленные одноклеточными водорослями, могут существовать лишь до тех пор, пока сквозь толщу воды способен проникать солнечный свет. Это глубина 150—200 метров. Гетеротрофы могут жить и в более глубоких слоях, в ночное время поднимаясь к поверхности для подкормки водорослями, а утром вновь уходя на обычную глубину, совершая при этом вертикальные миграции до 1 километра в сутки. В свою очередь, гетеротрофы, являющиеся консументами последующих порядков, обитающие ещё глубже, утром поднимаются до уровня обитания консументов I порядка, чтобы питаться ими.
Таким образом, мы видим, что в глубоких водоёмах, как правило, морях и океанах, существует такое понятие, как «пищевая лестница». Его смысл заключается в том, что органические вещества, которые создаются водорослями в поверхностных слоях земли, переносятся по пищевой цепочке до самого дна. Учитывая этот факт, можно считать обоснованным мнение некоторых экологов о том, что весь водоём можно считать единым биогеоценозом.
Детритные трофические взаимосвязи
Чтобы понять, что такое детритная пищевая цепь, нужно начать с самого понятия «детрит». Детрит — это совокупность остатков отмерших растений, трупов и конечных продуктов обмена животных.
Детритные цепи являются типичными для сообществ внутриконтинентальных вод, дна озер, имеющих большую глубину, и океанов, многие представители которых питаются именно детритом, образованным остатками мёртвых организмов из верхних слоев или случайно попавших в водоем из экологических систем, находящихся на суше, в виде, например, листового опада.
Донные экологические системы океанов и морей, где нет продуцентов ввиду отсутствия солнечного света, и вовсе могут существовать только за счёт детрита, общая масса которого в Мировом океане за календарный год может достигать сотни миллионов тонн.
Также детритные цепи распространены в лесах, где немалая часть ежегодного прироста биомассы продуцентов не может быть употреблена в пищу непосредственно первым звеном консументов. Поэтому она отмирает, образуя опад, который, в свою очередь, разлагается сапротрофами, а затем минерализуется редуцентами. Важную в роль в образовании детрита лесных сообществ играют грибы.
Гетеротрофы, которые питаются непосредственно детритом — это детритофаги. В наземных экологических системах к детритофагам относят некоторые виды членистоногих, в частности насекомых, а также кольчатых червей. Крупных детритофагов среди птиц (грифов, ворон) и млекопитающих (гиен) принято называть падальщиками.
В экологических системах вод основную массу детритофагов составляют водные насекомые и их личинки, а также некоторые представители ракообразных. Детритофаги могут служить пищей для более крупных гетеротрофов, которые также, в свою очередь, в дальнейшем могут стать пищей для консументов высших порядков.
Паразитические цепи
Трофические цепи начинаются со свободноживущего организма, на котором могут вести паразитический образ жизни паразиты I порядка, а на них уже могут паразитировать паразиты II порядка и т. д.
Пример: Круглый червь аскарида паразитирует на холоднокровных животных и является паразитом I порядка, но и она, в свою очередь, может быть хозяином для паразитирующих в её клетках одноклеточных эукариотов, паразитов II порядка, которые будут хозяевами для паразитов III порядка — бактерий, являющихся хозяевами, но уже для паразитов IV порядка — вирусов (бактериофагов).
Трофические уровни
Звенья пищевой цепочки иначе называют трофическими уровнями. По определению это группа организмов, которая занимает конкретное место в пищевой цепочке и представляющая для каждого из последующих уровней источник энергии — пищу.
Организмами I трофического уровня в пастбищных пищевых цепях являются первичные продуценты, автотрофы, то есть растения, и хемотрофы — бактерии, использующие энергию химических реакций для синтеза органических веществ. В детритных же системах автотрофы отсутствуют, а I трофический уровень детритной трофической цепи образует собственно детрит.
II трофический уровень пищевой цепочки формируют консументы I порядка — растительноядные организмы, называемые фитофагами. Сюда же относят высших и низших растений-паразитов, а также представителей зоопланктона, которые употребляют в пищу фитопланктон.
III трофический уровень образуют консументы II порядка — хищные животные — зоофаги, которые употребляют в качестве пищи фитофагов.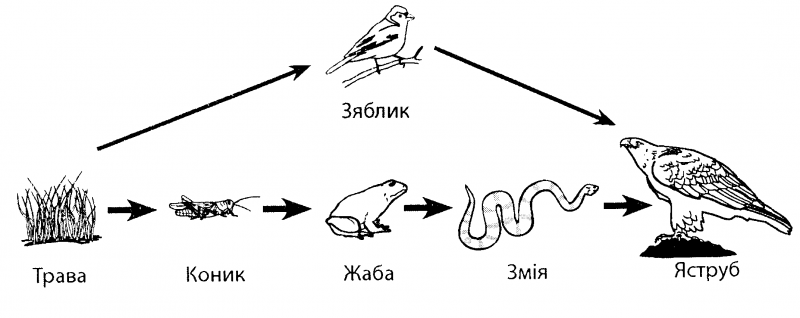 Это хищники I порядка — небольшие млекопитающие и птицы, основной рацион которых составляют насекомые. К этой группе также относят эндо- и эктопаразитов зоофагов, а также тех фитофагов, которые питаются растениями-паразитами.
Это хищники I порядка — небольшие млекопитающие и птицы, основной рацион которых составляют насекомые. К этой группе также относят эндо- и эктопаразитов зоофагов, а также тех фитофагов, которые питаются растениями-паразитами.
IV трофический уровень формируется консументами III порядка — более крупными хищниками II порядка, а также животными, которые паразитируют на хищниках I порядка.
Последний, V трофический уровень представлен организмами, которые потребляют мёртвые органические вещества и конечные продукты распада. Эти организмы называют деструкторами или редуцентами. Редуценты в основном представлены беспозвоночными животными, являющимися некро-, сапро- и копрофагами, использующими в пищу остатки, отходы и мёртвую органику. Также к этой группе относят растения-сапрофаги, которые разлагают листовой опад.
Ещё к уровню деструктуров относят гетеротрофные микроорганизмы, способные превращать органические вещества в неорганические (минеральные), образуя окончательные продукты — двуокись углерода и воду, которые возвращаются в экологическую систему и вновь вступают в природный круговорот веществ.
Значение пищевых взаимосвязей
- Изучение пищевых цепей помогает лучше понять взаимоотношения между разными организмами и проследить их взаимодействие между собой в окружающем мире.
- Благодаря наличию трофических цепей у исследователей есть возможность правильно определить механизм энергетических потоков и циркуляцию веществ в экосистеме.
- На каждый последующий уровень из предыдущего переходит максимум 15% энергии. Соответственно, продуцентов намного больше, чем консументов. А консументов намного больше, чем редуцентов. Это так называемая проблема биоусиления. Однако, несмотря на огромную конкуренцию, все организмы взаимосвязаны, и смерть всего лишь одного вида может воздействовать на все остальные и привести к совершенно непредсказуемым последствиям. Соответственно, пищевые цепи в экосистеме являются её важными составляющими, без которых она не сможет нормально функционировать.
Системы цитохрома Р450 – биологические варианты электронтранспортных цепей
Обзор
. 2007 март; 1770 (3): 330-44.
2007 март; 1770 (3): 330-44.
doi: 10.1016/j.bbagen.2006.07.017. Epub 2006 2 августа.
Франк Ханнеманн 1 , Андреас Бише, Керстин М. Юэн, Рита Бернхардт
принадлежность
- 1 FR 8.3-Биохимия, Саарландский университет, D-66041 Саарбрюккен, Германия.
- PMID: 16978787
- DOI: 10.1016/j.bbagen.2006.07.017
Обзор
Frank Hannemann et al. Биохим Биофиз Акта. 2007 9 марта0003
. 2007 март; 1770 (3): 330-44.
2007 март; 1770 (3): 330-44.
doi: 10.1016/j.bbagen.2006.07.017. Epub 2006 2 августа.
Авторы
Франк Ханнеманн 1 , Андреас Бише, Керстин М. Юэн, Рита Бернхардт
принадлежность
- 1 FR 8.3-Биохимия, Саарландский университет, D-66041 Саарбрюккен, Германия.
- PMID: 16978787
- DOI: 10.1016/j.bbagen.2006.07.017
Абстрактный
Цитохромы Р450 (Р450) представляют собой гемопротеины, кодируемые суперсемейством генов, почти повсеместно распространенных в различных организмах из всех биологических царств.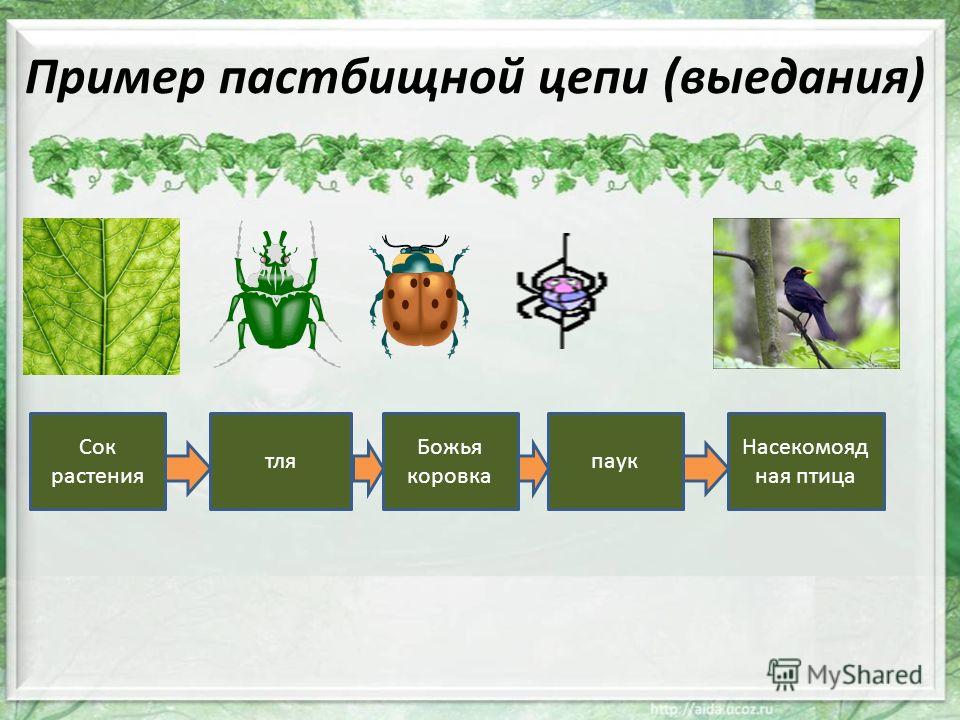 Реакции, осуществляемые P450, чрезвычайно разнообразны и способствуют биотрансформации лекарств, биоконверсии ксенобиотиков, биоактивации химических канцерогенов, биосинтезу физиологически важных соединений, таких как стероиды, жирные кислоты, эйкозаноиды, жирорастворимые витамины и желчные кислоты. , превращение алканов, терпенов и ароматических соединений, а также разложение гербицидов и инсектицидов. Цитохромы Р450 принадлежат к группе внешних монооксигеназ и, таким образом, получают необходимые электроны для расщепления кислорода и гидроксилирования субстрата от различных окислительно-восстановительных партнеров. Классические, а также недавно обнаруженные окислительно-восстановительные системы P450 собраны в этой статье и классифицированы в соответствии с их составом.
Реакции, осуществляемые P450, чрезвычайно разнообразны и способствуют биотрансформации лекарств, биоконверсии ксенобиотиков, биоактивации химических канцерогенов, биосинтезу физиологически важных соединений, таких как стероиды, жирные кислоты, эйкозаноиды, жирорастворимые витамины и желчные кислоты. , превращение алканов, терпенов и ароматических соединений, а также разложение гербицидов и инсектицидов. Цитохромы Р450 принадлежат к группе внешних монооксигеназ и, таким образом, получают необходимые электроны для расщепления кислорода и гидроксилирования субстрата от различных окислительно-восстановительных партнеров. Классические, а также недавно обнаруженные окислительно-восстановительные системы P450 собраны в этой статье и классифицированы в соответствии с их составом.
Похожие статьи
Цитохромы Р450 как универсальные биокатализаторы.
Бернхардт Р.
Бернхардт Р.
Дж Биотехнолог. 2006 25 июня; 124 (1): 128-45. doi: 10.1016/j.jbiotec.2006.01.026. Epub 2006 3 марта.
Дж Биотехнолог. 2006.
PMID: 16516322
Обзор.Активация кислорода монооксигеназой цитохрома Р450.
Хамдане Д., Чжан Х., Холленберг П. Хамдейн Д. и др. Фотосинтез Рез. 2008 г., октябрь-декабрь; 98 (1-3): 657-66. doi: 10.1007/s11120-008-9322-1. Epub 2008 4 июля. Фотосинтез Рез. 2008. PMID: 18600471 Бесплатная статья ЧВК. Обзор.
Цитохромы Р450 человека: проблемы и перспективы.
Гонсалес Ф.Дж. Гонсалес Ф.Дж. Trends Pharmacol Sci. 1992 сен; 13 (9): 346-52. дои: 10.1016/0165-6147(92)
-ч. Trends Pharmacol Sci. 1992. PMID: 1529480 Обзор.Изоферменты цитохрома Р450 в метаболизме эндо- и экзогенных соединений.
Вишневска А, Мазерска З. Вишневская А. и др. Постэпи Биохим. 2009;55(3):259-71. Постэпи Биохим. 2009. PMID: 19928582 Обзор. польский.
Цитохромы Р450 в биоактивации химических веществ.
Иоаннидес К., Льюис Д.Ф. Иоаннидес С. и др. Curr Top Med Chem. 2004;4(16):1767-88. дои: 10.2174/1568026043387188. Curr Top Med Chem. 2004. PMID: 15579107 Обзор.
 Бернхардт Р.
Дж Биотехнолог. 2006 25 июня; 124 (1): 128-45. doi: 10.1016/j.jbiotec.2006.01.026. Epub 2006 3 марта.
Дж Биотехнолог. 2006.
PMID: 16516322
Обзор.
Бернхардт Р.
Дж Биотехнолог. 2006 25 июня; 124 (1): 128-45. doi: 10.1016/j.jbiotec.2006.01.026. Epub 2006 3 марта.
Дж Биотехнолог. 2006.
PMID: 16516322
Обзор.
Посмотреть все похожие статьи
Цитируется
Роль фермента цитохрома P450 в коммуникации растительных микроорганизмов: в центре внимания виноградная лоза.
Минерди Д., Савой С., Саббатини П. Минерди Д. и соавт. Int J Mol Sci. 2023 28 февраля; 24 (5): 4695.
дои: 10.3390/ijms24054695.
Int J Mol Sci. 2023.
PMID: 366 Бесплатная статья ЧВК. Обзор.
Эволюция ферментов цитохрома P450 и их окислительно-восстановительных партнеров у архей.
Ngcobo PE, Nkosi BVZ, Chen W, Nelson DR, Syed K. Ngcobo PE и др. Int J Mol Sci. 2023 19 февраля;24(4):4161. дои: 10.3390/ijms24044161. Int J Mol Sci. 2023. PMID: 36835573 Бесплатная статья ЧВК.
Взаимное влияние ферментов цитохрома Р450 человека и УДФ-глюкуронозилтрансфераз на их соответствующую активность в рекомбинантных делящихся дрожжах.
Шарма С.С., Шарма С., Чжао Дж., Бурейк М. Шарма С.С. и др. Биомедицины. 2023 19 января; 11 (2): 281. doi: 10.3390/биомедицина11020281. Биомедицины. 2023. PMID: 36830817 Бесплатная статья ЧВК.
Предпочтительное для тканей рекрутирование цепей переноса электронов для катализируемого цитохромом P450 фенольного биосинтеза.
Чжао Х, Чжао Ю, Гоу М, Лю СиДжей. Чжао X и др. Научная реклама 2023 13 января; 9 (2): eade4389. doi: 10.1126/sciadv.ade4389. Epub 2023 11 января. Научная реклама 2023. PMID: 36630494 Бесплатная статья ЧВК.
Возможные механизмы взаимодействия между наркотиками и микотоксинами – возможно ли воздействие?
Лоотенс О., Вермеулен А., Крубельс С., Де Сэгер С., Ван Бокслаер Дж., Де Боевр М. Лутенс О. и др. Токсины (Базель). 2022 14 декабря; 14 (12): 873. doi: 10.3390/toxins14120873. Токсины (Базель). 2022. PMID: 36548770 Бесплатная статья ЧВК. Обзор.
 дои: 10.3390/ijms24054695.
Int J Mol Sci. 2023.
PMID: 36
дои: 10.3390/ijms24054695.
Int J Mol Sci. 2023.
PMID: 36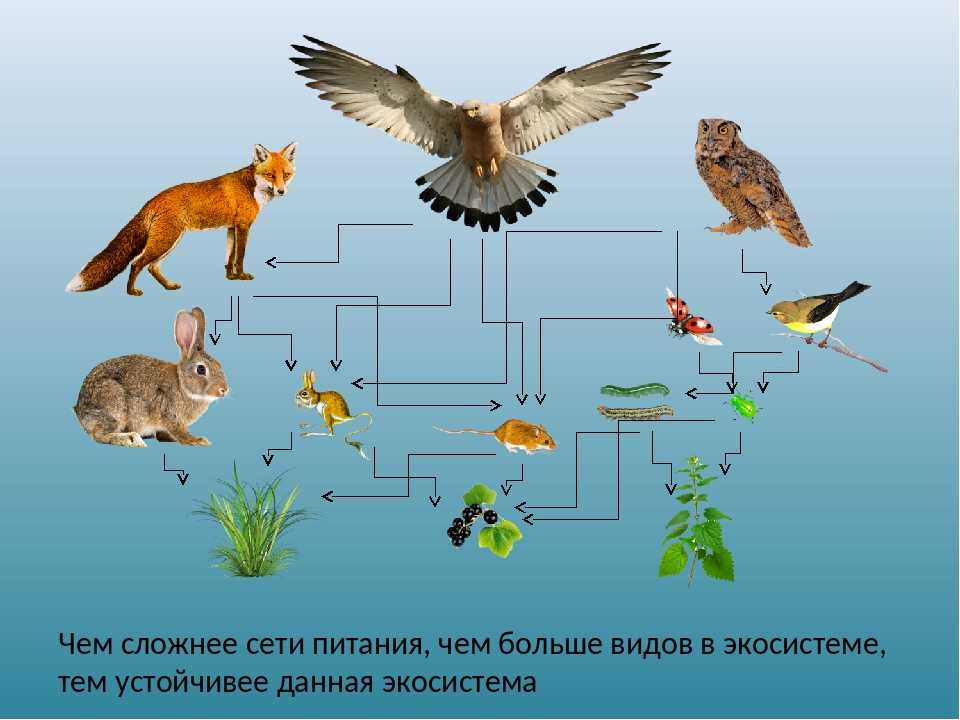
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Поиски биологических источников длинноцепочечных алкилдиолов в западной тропической части Северной Атлантики
Агата, С.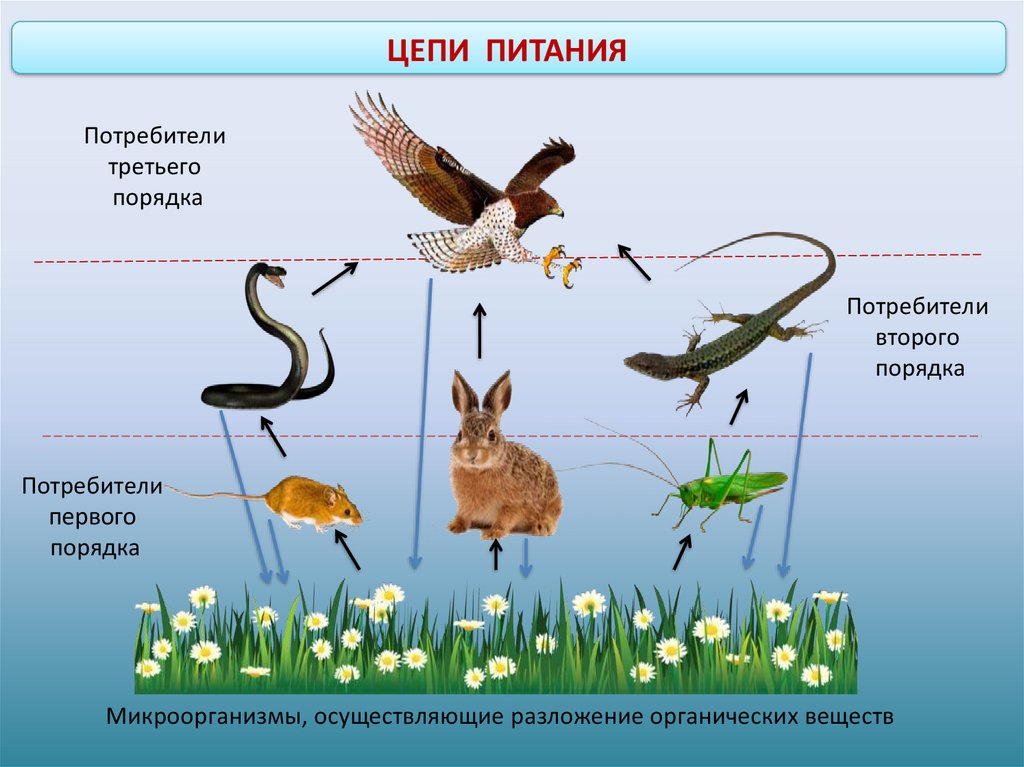 , Штрудер-Кипке, М.С., и Беран, А.: Морфологические и генетические
изменчивость морской планктонной инфузории Laboea strobila Lohmann, 1908 (Ciliophora,
Oligotrichia), с заметками об его онтогенезе, J. Eukaryot. микробиол., 51,
267–281, https://doi.org/10.1111/j.1550-7408.2004.tb00567.x, 2004.
, Штрудер-Кипке, М.С., и Беран, А.: Морфологические и генетические
изменчивость морской планктонной инфузории Laboea strobila Lohmann, 1908 (Ciliophora,
Oligotrichia), с заметками об его онтогенезе, J. Eukaryot. микробиол., 51,
267–281, https://doi.org/10.1111/j.1550-7408.2004.tb00567.x, 2004.
Альтшул С.Ф., Гиш В., Миллер В., Майерс Э.В. и Липман Д.Дж. : Инструмент для поиска основного локального выравнивания, J. Mol. био., 215, 403–410, https://doi.org/10.1016/S0022-2836(05)80360-2, 1990.
Андерсен Р.А., Бретт Р.В., Поттер Д. и Секстон Дж.П.: Филогенез
Eustigmatophyceae на основе 18S рДНК с акцентом на Nannochloropsis , Protist, 149, 61–74, https://doi.org/10.1016/S1434-4610(98)70010-0, 1998. CPD, Besseling, M.,
Дорхаут, Д., Синнинге Дамсте, Дж. С., и Схоутен, С.: C 5 гликолипиды гетероцистных цианобактерий отслеживают численность симбионтов в диатомовых водорослях Hemiaulus hauckii в тропической части Северной Атлантики, Biogeosciences, 15, 1229–1241, https://doi.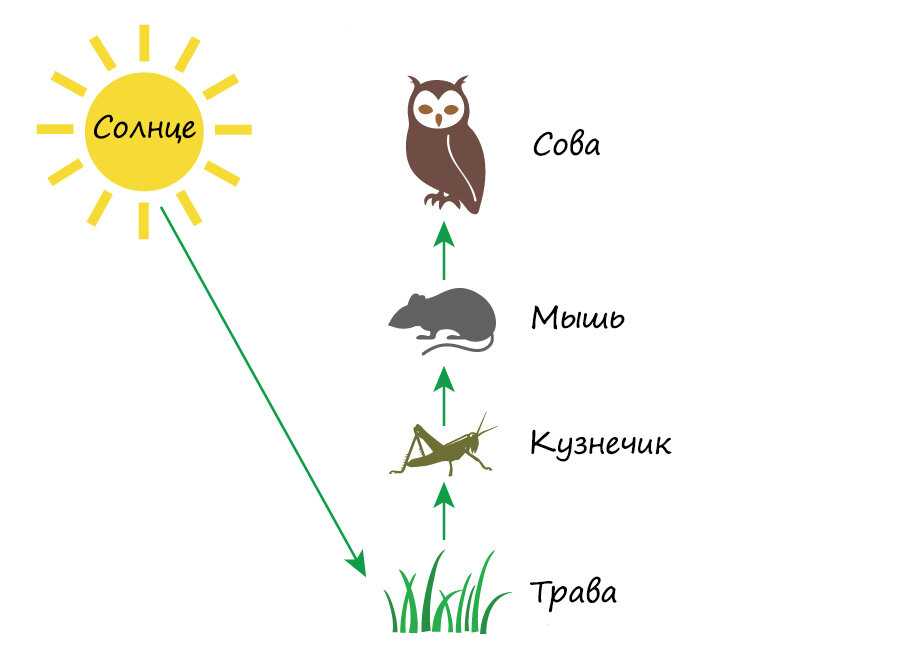 org/10.5194/bg-15-1229-2018, 2018.
org/10.5194/bg-15-1229-2018, 2018.
Balzano, S., Pancost, R.D., Lloyd , Дж. Р., и Стэтхэм, П. Дж.: Изменения в состав жирных кислот в разлагающихся агрегатах водорослей, Mar. Chem., 124, 2–13, https://doi.org/10.1016/j.marchem.2010.11.001, 2011.
Balzano, S., Abs, E., and Leterme, S.C.: Разнообразие протистов в зависимости от солености градиент в прибрежной лагуне, Aquat. микроб. экол., 74, 263–277, https://doi.org/10.3354/ame01740, 2015 г.
Бальзано С., Вильянуэва Л., де Бар М., Синнинге Дамсте Дж. С. и Схоутен, С.: Влияние условий культивирования на численность и состав длинноцепочечных алкилдиолов у видов рода Наннохлоропсис , Орг. геохим., 108, 9–17, https://doi.org/10.1016/j.orggeochem.2017.02.006, 2017.
Бенджамини, Ю. и Хохберг, Ю.: Контроль уровня ложных открытий. А практичный и мощный подход к множественному тестированию., J. Roy. Стат. соц. Б мет., 57, 289–300, 1995.
Бергеш, М., Одебрехт, К., и Моструп, О.: Нанофлагелляты из прибрежных
воды южной Бразилии (32 град. ю.ш.), бот. 51 марта, 35–50,
https://doi.org/10.1515/bot.2008.003, 2008.
ю.ш.), бот. 51 марта, 35–50,
https://doi.org/10.1515/bot.2008.003, 2008.
Buckles, L.K., Villanueva, L., Weijers, JWH, Verschuren, D., и Sinninghe Damste, JS: Связывание изопреноидального GDGT мембранного липида распределения с обилием генов окисляющих аммиак Thaumarchaeota и некультурные группы кранархеотов в толще воды тропического озера (озеро Чалла, Восточная Африка), окр. микробиол., 15, 2445–2462, https://doi.org/10.1111/1462-2920.12118, 2013.
Buschhaus, C., Peng, C., and Jetter, R.: 1,2- и 1,3-бифункциональные соединения кутикулярного воска Cosmos bipinnatus лепестки, Фитохимия, 91, 249–256, https://doi.org/10.1016/j.phytochem.2012.07.018, 2013.
Caporaso, JG, Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F.
Д., Костелло, Е.К., Фиерер, Н., Пена, А.Г., Гудрич, Дж.К., Гордон, Дж.
И., Хаттли Г. А., Келли С. Т., Найтс Д., Кениг Дж. Э., Лей Р. Э.,
Лозупоне, К.А., Макдональд, Д., Мюгге, Б.Д., Пиррунг, М., Ридер, Дж.,
Севински Дж. Р., Тамбо П. Дж., Уолтерс В.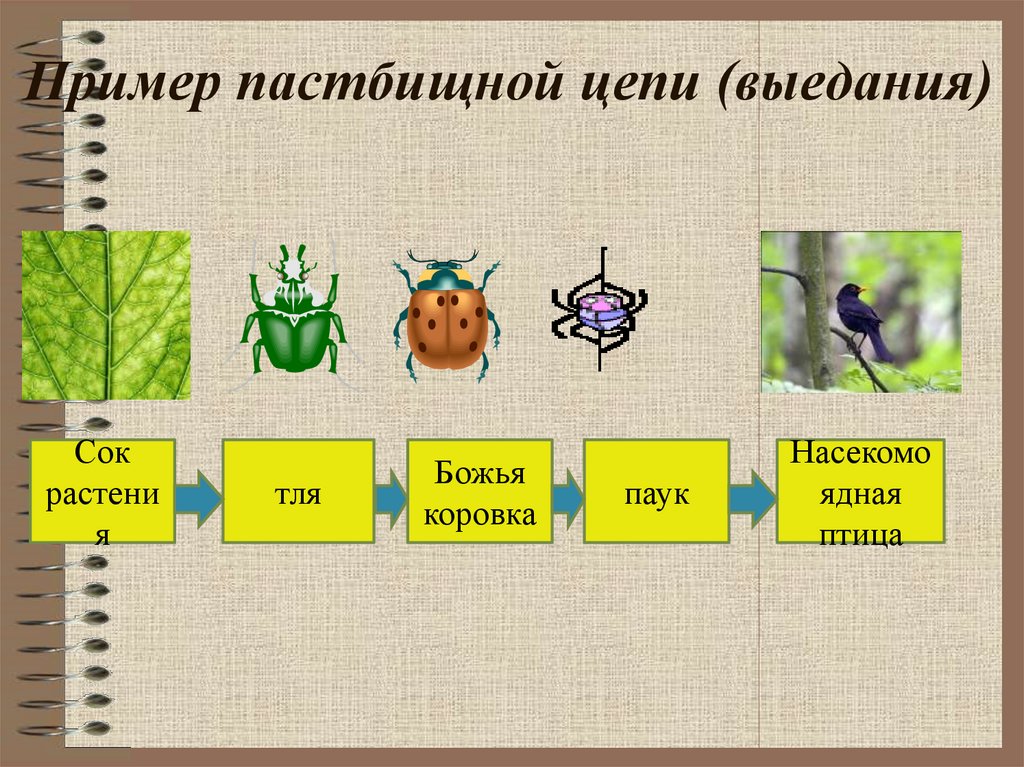 А., Видманн Дж., Яцуненко Г.
Т., Заневельд Дж. и Найт Р.: QIIME позволяет анализировать высокопроизводительные
данные секвенирования сообщества, Nat. Методы, 7, 335–336,
https://doi.org/10.1038/nmeth.f.303, 2010 г.
А., Видманн Дж., Яцуненко Г.
Т., Заневельд Дж. и Найт Р.: QIIME позволяет анализировать высокопроизводительные
данные секвенирования сообщества, Nat. Методы, 7, 335–336,
https://doi.org/10.1038/nmeth.f.303, 2010 г.
Кристаки У., Кормас К.А., Генитсарис С., Жорж К., Симе-Нгандо Т., Вискоглиози, Э., и Монши, С.: Зимне-летняя сукцессия одноклеточных эукариоты в мезоэвтрофной прибрежной системе, Microb. Экол., 67, 13–23, https://doi.org/10.1007/s00248-013-0290-4, 2014.
Comeau, A.M., Li, WKW, Tremblay, J.-E., Carmack, E.C., and Lovejoy, C.: Структура микробного сообщества Северного Ледовитого океана до и после 2007 г. рекордный минимум морского льда, Plos One, 6, e27492, https://doi.org/10.1371/journal.pone.0027492, 2011.
де Бар, М. В., Дорхаут, Д. Дж. К., Хопманс, Э. К., Рампен, С. В., Синнингхе
Дамсте, Дж. С., и Схоутен, С.: Ограничения на применение длинных
прокси цепных диолов на иберийско-атлантической окраине, Org. Геохим., 101, оф.
184–195, https://doi.org/10.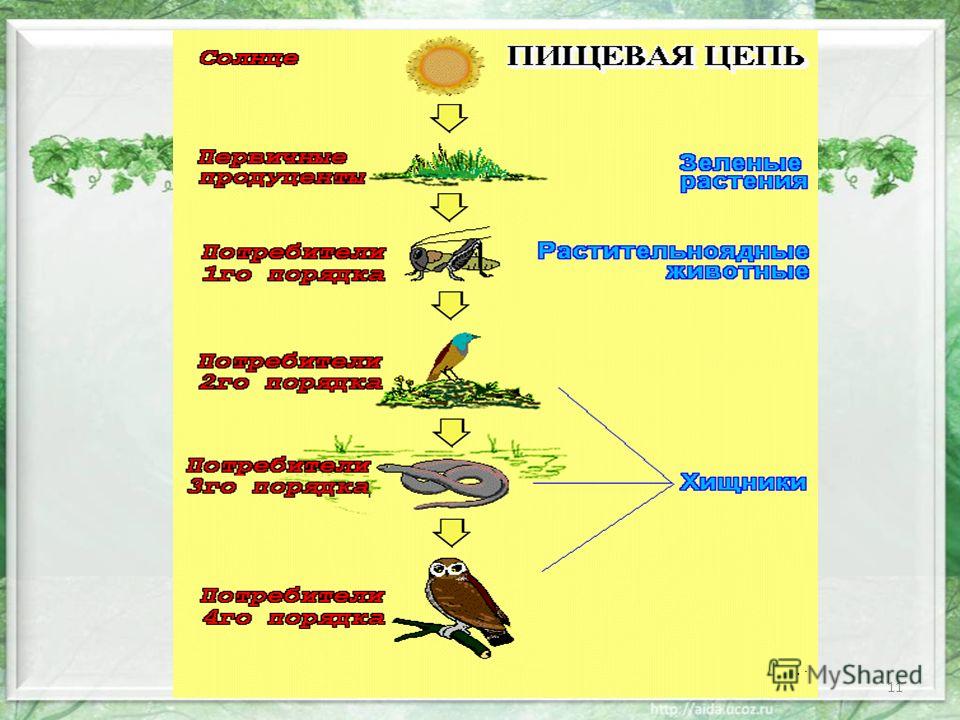 1016/j.orggeochem.2016.09.005, 2016.
1016/j.orggeochem.2016.09.005, 2016.
de Leeuw, J.W., Irene, W., Rijpstra, C., and Schenck, P.A.: Возникновение и идентификация C 30 , C 31 и C 32 алкан-1, 15-диолов и алкан-15-он-1-олы в отложениях Черного моря пачки I и пачки II // Геохим. Космохим. Ак., 45, 2281–2285, https://doi.org/10.1016/0016-7037(81)
-6, 1981.de Leeuw, J.W., Versteegh, GJM, and van Bergen, P.F.: Биомакромолекулы водорослей и растений и их ископаемые аналоги, Растение Ecol., 182, 209–233, https://doi.org/10.1007/s11258-005-9027-x, 2006.
Деренн, С. и Ларжо, К.: Обзор некоторых важных семейств тугоплавкие макромолекулы: состав, происхождение и судьба в почвах и осадки, Почвоведение, 166, 833–847, https://doi.org/10.1097/00010694-200111000-00008, 2001.
де Варгас, К., Аудик, С., Генри, Н., Десель, Дж., Маэ, Ф., Логарес, Р. ,
Лара, Э., Берни, К., Ле Беско, Н., Проберт, И., Кармайкл, М., Пулен,
Дж., Ромак С., Колин С., Аури Дж.-М., Биттнер Л. , Шаффрон С., Данторн,
М., Энгелен С., Флегонтова О., Гуиди Л., Хорак А., Жайон О.,
Лима-Мендес Г., Лукеш Дж., Мальвия С., Морар Р., Мюлот М., Скалько,
Э., Сиано Р., Винсент Ф., Зингоне А., Димье К., Пичераль М., Сирсон,
С., Канделс-Льюис С., Ачинас С.Г., Борк П., Боулер С., Горский Г.,
Гримсли Н., Хингэмп П., Юдикон Д., Нот Ф., Огата Х., Песант С.,
Раес Дж., Серацки М.Е., Спейч С., Штемманн Л., Сунагава С.,
Вайссенбах Дж., Винкер П. и Карсенти Э.: Эукариотический планктон.
разнообразие в освещенном солнцем океане, Science, 348, 1261605, https://doi.org/10.1126/science.1261605, 2015.
, Шаффрон С., Данторн,
М., Энгелен С., Флегонтова О., Гуиди Л., Хорак А., Жайон О.,
Лима-Мендес Г., Лукеш Дж., Мальвия С., Морар Р., Мюлот М., Скалько,
Э., Сиано Р., Винсент Ф., Зингоне А., Димье К., Пичераль М., Сирсон,
С., Канделс-Льюис С., Ачинас С.Г., Борк П., Боулер С., Горский Г.,
Гримсли Н., Хингэмп П., Юдикон Д., Нот Ф., Огата Х., Песант С.,
Раес Дж., Серацки М.Е., Спейч С., Штемманн Л., Сунагава С.,
Вайссенбах Дж., Винкер П. и Карсенти Э.: Эукариотический планктон.
разнообразие в освещенном солнцем океане, Science, 348, 1261605, https://doi.org/10.1126/science.1261605, 2015.
Dixon, P.: VEGAN, пакет функций R для экологии сообщества, J. Veg. Sci., 14, 927–930, https://doi.org/10.1111/j.1654-1103.2003.tb02228.x, 2003.
Эдгар, Р. К.: Поиск и кластеризация на порядки быстрее, чем BLAST, Биоинформатика, 26, 2460–2461, https://doi.org/10.1093/bioinformatics/btq461, 2010.
Эдгар, Р. К., Хаас, Б. Дж., Клементе, Дж. К., Айва, К., и Найт, Р.:
UCHIME повышает чувствительность и скорость обнаружения химер, биоинформатики,
27, 2194–2200, https://doi. org/10.1093/bioinformatics/btr381, 2011.
org/10.1093/bioinformatics/btr381, 2011.
Эгге, Э. С., Йоханнессен, Т. В., Андерсен, Т., Эйкрем, В., Биттнер, Л., Ларсен А., Сандаа Р.-А. и Эдвардсен Б.: Сезонное разнообразие и динамика гаптофитов в Скагерраке, Норвегия, исследованная высокопроизводительное секвенирование, мол. Экол., 24, 3026–3042, https://doi.org/10.1111/mec.13160, 2015.
Фоли, К.П. и Фоли, М.В.: Наблюдения за разнообразием и экологией пресной воды Наннохлоропсис (Eustigmatophyceae), с описанием новых таксонов, Protist, 158, 325–336, https://doi.org/10.1016/j.protis.2007.03.003, 2007.
Гелин Ф., Бугерс И., Норделоос А.А. Ригман, Р., и Де Леу, Дж. В.: Устойчивые биомакромолекулы в морских микроводоросли классов Eustigmatophyceae и Chlorophyceae: геохимические последствия, орг. Геохим., 26, 659–675, https://doi.org/10.1016/s0146-6380(97)00035-1, 1997а.
Гелин, Ф., Фолькман, Дж. К., де Лиу, Дж. В., и Синнинге Дамсте, Дж. С.:
Среднецепочечные гидроксидлинноцепочечные жирные кислоты микроводорослей рода Наннохлоропсис , Фитохимия, 45, 641–646,
https://doi. org/10.1016/s0031-9422(97)00068-x, 1997b.
org/10.1016/s0031-9422(97)00068-x, 1997b.
Гросси В., Блоккер П. и Синнинге Дамсте Дж. С.: Анаэробные биодеградация липидов морской микроводоросли Nannochloropsis salina , Org. Геохим., 32, оф. 795–808, https://doi.org/10.1016/s0146-6380(01)00040-7, 2001.
Guillou, L., Viprey, M., Chambouvet, A., Welsh, R.M., Kirkham, A.R. , Массана, Р., Сканлан, Д. Дж., и Уорден, А. З.: Широкое распространение и генетическое разнообразие морских паразитоидов, принадлежащих Syndiniales (Альвеолата), Окружающая среда. микробиол., 10, 3349–3365, https://doi.org/10.1111/j.1462-2920.2008.01731.x, 2008.
Guillou, L., Bachar, D., Audic, S., Bass, D., Berney, C., Bittner , Л.,
Бутте, К., Бурго, Г., де Варгас, К., Десель, Дж., дель Кампо, Дж., Долан,
Дж. Р., Данторн М., Эдвардсен Б., Хольцманн М., Коойстра В. Х. С. Ф.,
Лара, Э., Ле Беско, Н., Логарес, Р., Маэ, Ф., Массана, Р., Монтрезор,
М., Морар Р., Нот Ф., Павловски Дж., Проберт И., Соваде А.-Л., Сиано,
Р., Стоек Т., Воло Д.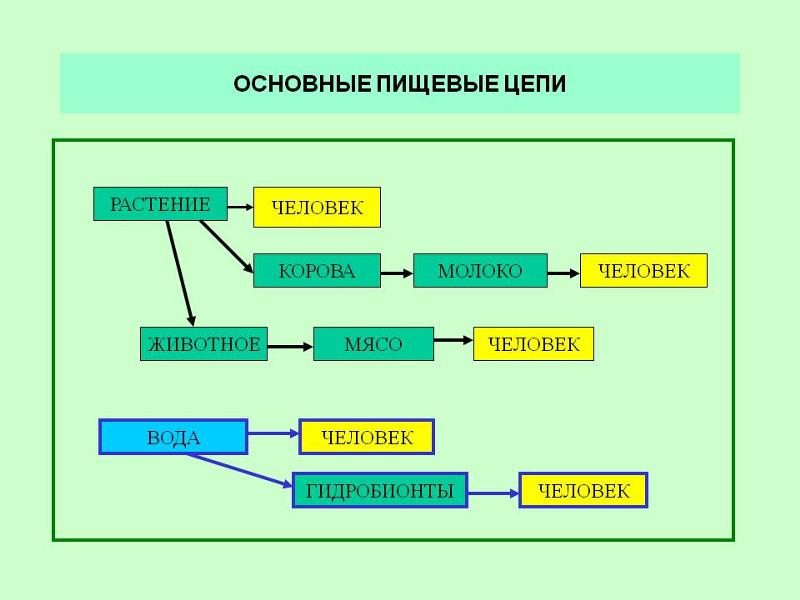 , Циммерманн П. и Кристен Р.: Протист
Ribosomal Reference database (PR2): каталог одноклеточных эукариот Small
Последовательности субъединиц рРНК с тщательно подобранной таксономией, Nucleic Acid Res., 41,
597–604, https://doi.org/10.1093/nar/gks1160, 2013.
, Циммерманн П. и Кристен Р.: Протист
Ribosomal Reference database (PR2): каталог одноклеточных эукариот Small
Последовательности субъединиц рРНК с тщательно подобранной таксономией, Nucleic Acid Res., 41,
597–604, https://doi.org/10.1093/nar/gks1160, 2013.
Джеттер, Р. и Ридерер, М.: Алкандиолы с длинной цепью, кетоальдегиды, кетоспирты и кетоалкиловые эфиры в кутикулярном воске Osmunda regalis вайи, Фитохимия, 52, 907–915, https://doi.org/10.1016/s0031-9422(99)00309-x, 1999.
Цзян, С. К., Олеари, Т., Фолькман, Дж. К., Чжан, Х. З., Цзя, Р. Ф., Ю, С. Х., Ван Ю., Луан З.Ф., Сунь З.К. и Цзян Р.Х.: Истоки и смоделированное термическое изменение стеролов и кетоспиртов в глубоководных морских отложения Окинавского прогиба, орг. Геохим., 21, 415–422, https://doi.org/10.1016/0146-6380(94)
-8, 1994. Конинг, Э., ван Иперен, Дж. М., ван Раафорст, В., Хелдер, В., Браммер, Г. Дж.
А., и ван Веринг, Т. С. Е.: Избирательное сохранение
диатомовые водоросли, указывающие на апвеллинг, в отложениях у берегов Сомали, на северо-западе Индийского океана,
Глубоководные рез. Пт. I, 48, 2473–2495,
https://doi.org/10.1016/s0967-0637(01)00019-x, 2001.
Пт. I, 48, 2473–2495,
https://doi.org/10.1016/s0967-0637(01)00019-x, 2001.
Ласситер, А. М., Вилкерсон, Ф. П., Дагдейл, Р. К., и Хог, В. Э.: Комплексы фитопланктона в прибрежной зоне апвеллинга CoOP-WEST, Deep-Sea Рез. Пт. II, 53, 3063–3077, https://doi.org/10.1016/j.dsr2.2006.07.013, 2006 г.
Латто, Дж., Дорхаут, Д., Шульц, Х., Кастанеда, И. С., Шефус, Э., Синнинге Дамсте, Дж. С., и Схоутен, С.: Алкан-1,15-диол C 32 как показатель позднечетвертичного периода. приток рек на прибрежные окраины, Clim. Прошлое, 13, 1049–1061, https://doi.org/10.5194/cp-13-1049-2017, 2017a.
Латто, Дж., Ким, Дж. Х., Де Йонге, К., Целль, К., Синнинге Дамсте, Дж. С., и Schouten, S.: Алкан-1,15-диол C 32 в качестве индикатора для речных вход в прибрежные моря, Геохим. Космохим. Ак., 202, 146–158, https://doi.org/10.1016/j.gca.2016.12.030, 2017b.
Лежандр П. и Галлахер Э. Д.: Экологически значимые преобразования
для ординации данных о видах, Oecologia, 129, 271–280,
https://doi. org/10.1007/s004420100716, 2001.
org/10.1007/s004420100716, 2001.
Логарес Р., Аудик С., Сантини С., Пернис М.С., де Варгас С. и Массана, Р.: Модели разнообразия и активность некультивируемых морских животных. гетеротрофные жгутиконосцы, обнаруженные с помощью пиросеквенирования, ISME J., 6, 1823–1833, https://doi.org/10.1038/ismej.2012.36, 2012.
Лопес душ Сантуш, А., Гурвиль, П., Трагин, М., Ноэль, М.-Х., Десель, Дж. ., Ромак С. и Воло Д.: Разнообразие и океаническое распространение prasinophytes клада VII, доминирующая группа зеленых водорослей в океане. воды, ISME J., 11, 512–528, https://doi.org/10.1038/ismej.2016.120, 2016.
Мао, С.Ю., Чжу, X.В., Ву, Н.Ю., Цзя, Г.Д., Сунь, Ю.Г., Гуань, Х.Х., и Ву, Д. Д.: Спиртовые соединения в Azolla imbricata и потенциальный источник значение для морских отложений, Sci. China Earth Science, 60, 348–359, https://doi.org/10.1007/s11430-016-5177-6, 2017.
Мари Д., Саймон Н. и Воло Д.: Подсчет клеток фитопланктона по потоку
цитометрия., в: Методы культивирования водорослей, под редакцией: RA, A.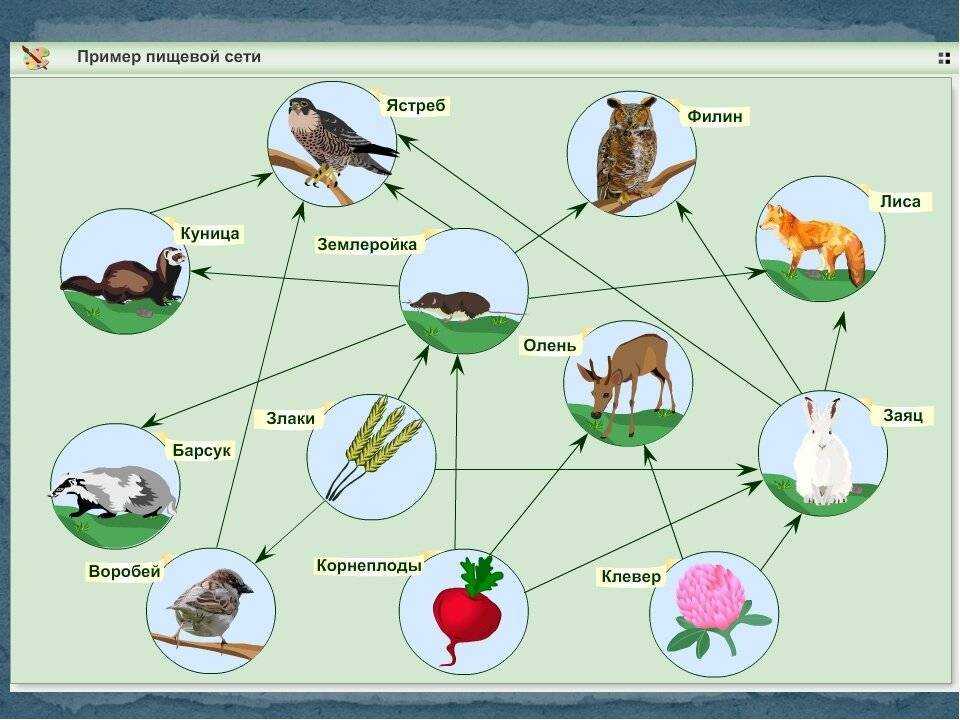 , Elsevier
Academic Press, Burlington, MA, 253–268, 2005.
, Elsevier
Academic Press, Burlington, MA, 253–268, 2005.
Massana, R., Gobet, A., Audic, S., Bass, D., Bittner, L., Boutte, C., Шамбуве, А., Кристен, Р., Клавери, Дж. М., Десель, Дж., Долан, Дж. Р., Дунторн М., Эдвардсен Б., Форн И., Форстер Д., Гийу Л., Жайон, О., Коойстра В., Логарес Р., Маэ Ф., Нот Ф., Огата Х., Павловски Дж., Пернис, М. К., Проберт, И., Ромак, С., Ричардс, Т., Сантини, С., Шальчиан-Табризи К., Сиано Р., Саймон Н., Стоек Т., Воло Д., Зингоне, А., и де Варгас, К.: Разнообразие морских простейших в прибрежных районах Европы. воды и отложения, выявленные высокопроизводительным секвенированием, Environ. микробиол., 17, 4035–4049., https://doi.org/10.1111/1462-2920.12955, 2015.
Мойта, М. Т., Оливейра, П. Б., Мендес, Дж. К., и Пальма, А. С.: Распространение хлорофилла а и Gymnodinium catenatum, связанного с прибрежным апвеллингом шлейфы у берегов центральной Португалии, Acta Oecol., 24, S125–S132, https://doi.org/10.1016/s1146-609x(03)00011-0, 2003.
Моллери, Г. С. Ф., Ново, Э., и Кампель, М.: Пространственно-временная изменчивость
Шлейф реки Амазонки на основе цвета океана со спутника, продолжение. Полка рез., 30,
342–352, https://doi.org/10.1016/j.csr.2009.11.015, 2010.
С. Ф., Ново, Э., и Кампель, М.: Пространственно-временная изменчивость
Шлейф реки Амазонки на основе цвета океана со спутника, продолжение. Полка рез., 30,
342–352, https://doi.org/10.1016/j.csr.2009.11.015, 2010.
Нильсен, К.М., Джонсен, П.Дж., Бенсассон, Д., и Даффонкио, Д.: Выпуск и устойчивость внеклеточной ДНК в окружающей среде, Экологическая Biosafety Research, 6, 37–53, https://doi.org/10.1051/ebr:2007031, 2007.
Not, F., del Campo, J., Balague, V., de Vargas, C., и Массана, Р.: Новый Insights in the Diversity of Marine Picoeukaryotes, Plos One, 4, e7143, https://doi.org/10.1371/journal.pone.0007143, 2009.
Питчер А., Вильянуэва Л., Хопманс Э. К., Схоутен С., Рейхарт Г. Дж., и Sinninghe Damste, JS: Нишевое разделение архей, окисляющих аммиак. и анаммокс-бактерии в зоне кислородного минимума Аравийского моря, ISME J., 5, 1896–1904, https://doi.org/10.1038/ismej.2011.60, 2011.
Планк, Дж., Маттиоли, Э., Питте, Б., Саймон, Л., и Гросси, В.:
Изменения продуктивности и температуры поверхности моря, зарегистрированные в конце
Эоцен-ранний олигоцен на участке DSDP 511 (Южная Атлантика), Палеогеогр.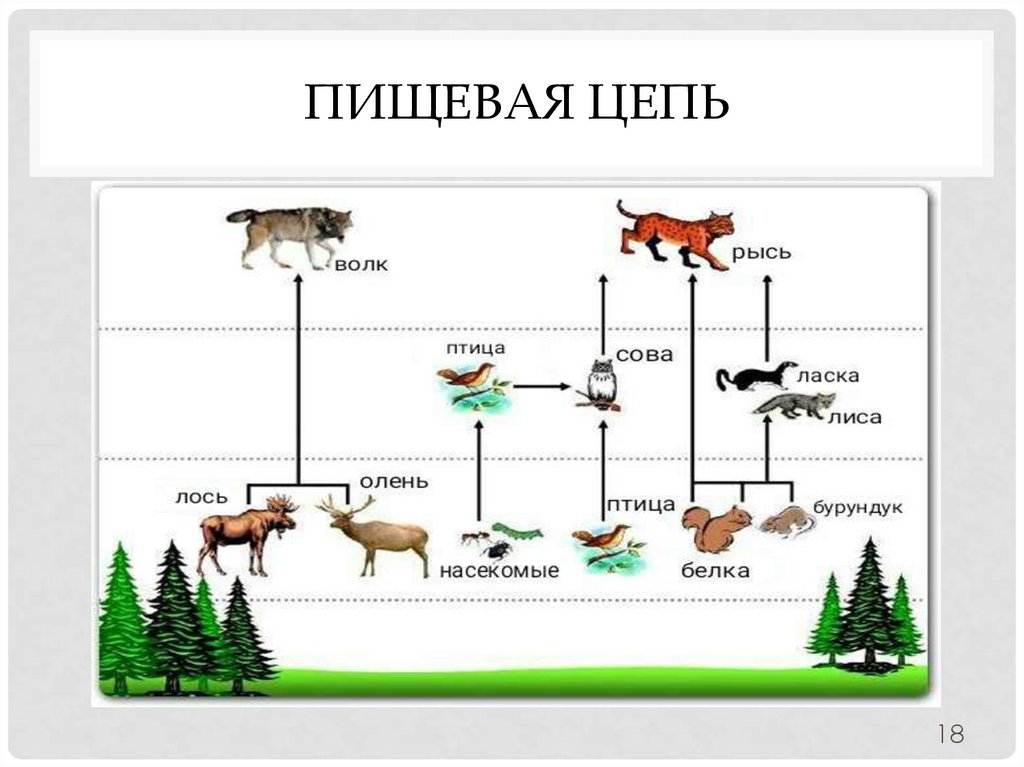 Palaeocl., 407, 34–44, https://doi.org/10.1016/j.palaeo.2014.04.016, 2014.
Соединения и метилразветвленные алканы в кутикулярных восках Triticum
аестивум сорт. Вифлеем, Плос Один, 11, 25,
https://doi.org/10.1371/journal.pone.0165827, 2016 г.
Palaeocl., 407, 34–44, https://doi.org/10.1016/j.palaeo.2014.04.016, 2014.
Соединения и метилразветвленные алканы в кутикулярных восках Triticum
аестивум сорт. Вифлеем, Плос Один, 11, 25,
https://doi.org/10.1371/journal.pone.0165827, 2016 г.
Rampen, S.W., Schouten, S., Wakeham, S.G., and Sinninghe Damste, J.S.: Сезонная и пространственная изменчивость источников и потоков длинноцепочечных диолов и среднецепочечные гидроксиметилалканоаты в Аравийском море, Org. Геохим., 38, 165–179, https://doi.org/10.1016/j.orggepchem.2006.10.008, 2007.
Rampen, S.W., Schouten, S., Koning, E., Brummer, G.-J. А. и Синнинге Дамсте, Дж. С.: 90-тысячелетний апвеллинг из северо-западной части Индии. Океан с использованием нового индекса длинноцепочечных диолов, Планета Земля. наук Летт., 276, 207–213, https://doi.org/10.1016/j.epsl.2008.09.022, 2008.
Rampen, S.W., Schouten, S., Schefuss, E., and Sinninghe Damste, J.S.:
Влияние температуры на диол с длинной цепью и гидроксиметил со средней длиной цепи
состав алканоатов в диатомовых водорослях Proboscia : результаты культуральных и полевых исследований, Org. Геохим., 40, 1124–1131, https://doi.org/10.1016/j.orggeochem.2009.08.005,
2009.
Геохим., 40, 1124–1131, https://doi.org/10.1016/j.orggeochem.2009.08.005,
2009.
Rampen, S.W., Schouten, S., and Sinninghe Damsté, J.S.: Возникновение длинноцепочечные 1,14-диолы в Apedinella radians , Org. Геохим., 42, 572–574, https://doi.org/10.1016/j.orggeochem.2011.03.009, 2011.
Rampen, S.W., Willmott, V., Kim, J.-H., Uliana, E., Mollenhauer, G., Шефус, Э., Синнинге Дамсте, Дж. С., и Схоутен, С.: Длинная цепь 1,13- и 1,15-диолы как потенциальный показатель палеотемпературы реконструкция, Геохим. Космохим. Ак., 84, 204–216, https://doi.org/10.1016/j.gca.2012.01.024, 2012.
Rampen, S.W., Datema, M., Rodrigo-Gamiz, M., Schouten, S., Reichart, G.-J. , и Синнинге Дамсте, Дж. С.: Источники и прокси-потенциал длинной цепи алкилдиолы в озерных средах // Геохим. Космохим. Ак., 144, 59–71, https://doi.org/10.1016/j.gca.2014.08.033, 2014а.
Рампен С.В., Уиллмотт В., Ким Дж.-Х., Родриго-Гамиз М., Ульяна Э.,
Молленхауэр, Г., Шефус, Э., Синнинге Дамсте, Дж.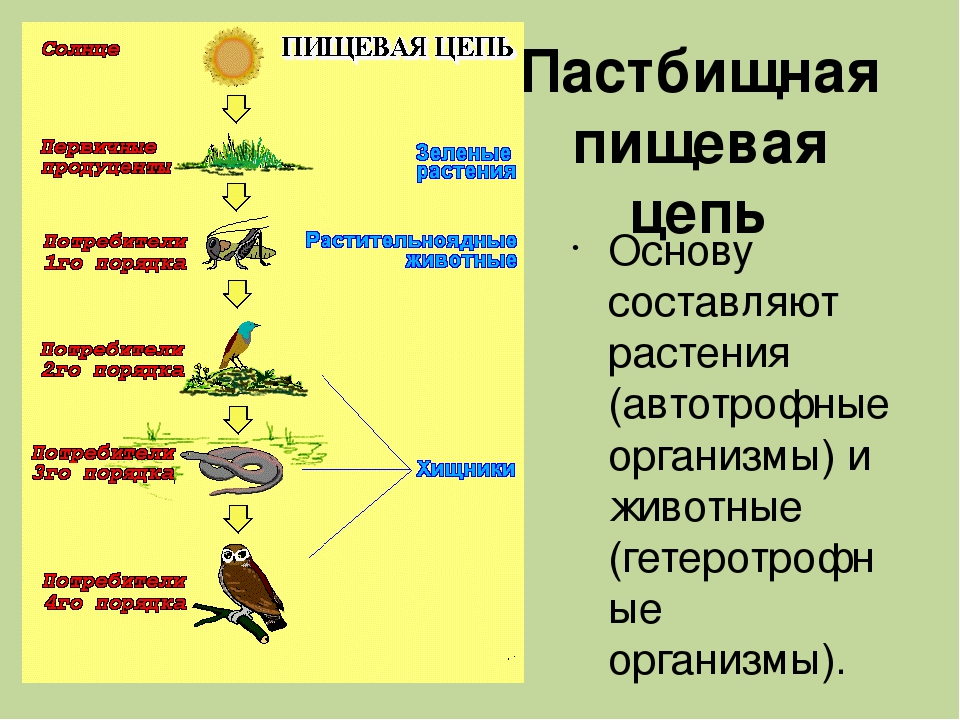 С., и Схоутен,
S.: Оценка длинноцепочечных 1,14-алкилдиолов в морских отложениях как
индикаторы апвеллинга и температуры, Орг. геохим., 76, 39–47,
https://doi.org/10.1016/j.orggeochem.2014.07.012, 2014б.
С., и Схоутен,
S.: Оценка длинноцепочечных 1,14-алкилдиолов в морских отложениях как
индикаторы апвеллинга и температуры, Орг. геохим., 76, 39–47,
https://doi.org/10.1016/j.orggeochem.2014.07.012, 2014б.
Райхе С., Рампен С., Дорхаут Д., Синнинге Дамсте Дж. С. и Схоутен S.: Воздействие кислорода на длинноцепочечные алкилдиолы и длинноцепочечные индекс цепных диолов (LDI) — долгосрочное инкубационное исследование, Biogeosciences Discuss., обзор, 2018 г.
Родриго-Гамис, М., Рампен, С.В., де Хаас, Х., Баас, М., Схоутен, С., и Синнинге Дамсте, Дж. С.: Ограничения применимости органических прокси температуры U37K′, TEX 86 и LDI в приполярном регионе вокруг Исландии, Biogeosciences, 12, 6573–6590, https://doi.org/10.5194/bg-12-6573-2015, 2015. классификация и разнообразие хореотрихий и олиготрихий (Ciliophora, Spirotrichea), мол. Филогенет. Эволюция, 112, 12–22, https://doi.org/10.1016/j.ympev.2017.03.010, 2017.
Шлитцер, Р.: Интерактивный анализ и визуализация геолого-геофизических данных
с Ocean Data View, Comput.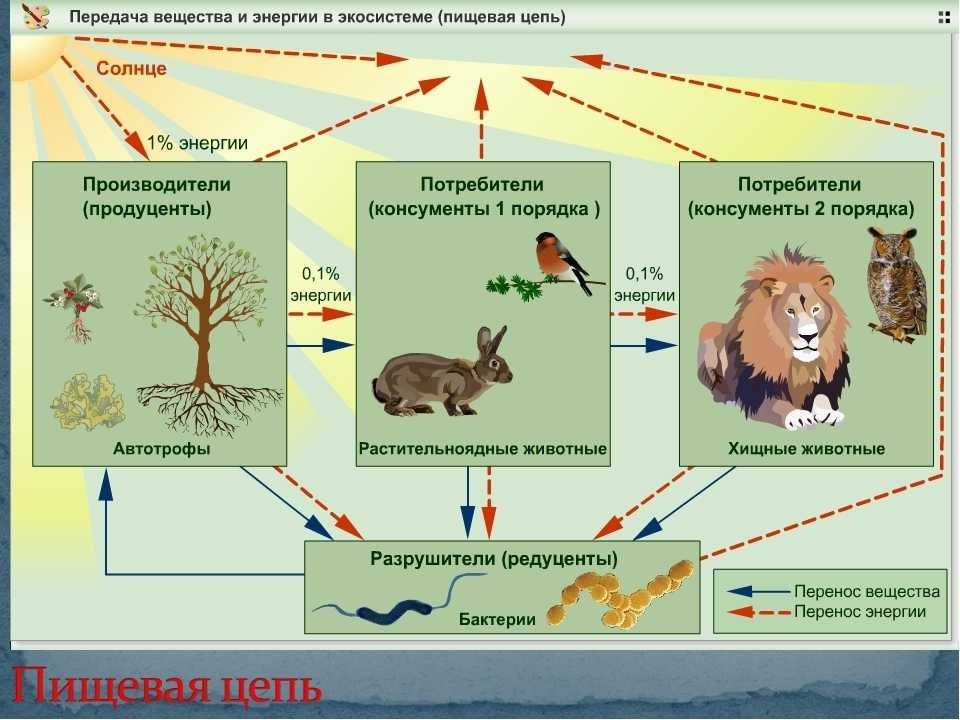 Geosci., 28, 1211–1218,
https://doi.org/10.1016/s0098-3004(02)00040-7, 2002.
Geosci., 28, 1211–1218,
https://doi.org/10.1016/s0098-3004(02)00040-7, 2002.
Шольц, М. Дж., Вайс, Т. Л., Джинкерсон, Р. Э., Цзин, Дж., Рот, Р., Гуденаф, У., Позевиц, М.С., и Геркен, Х.Г.: Ультраструктура и состав клеточной стенки Nannochloropsis gaditana эукариот. Ячейка, 13, 1450–1464, https://doi.org/10.1128/ec.00183-14, 2014.
Сеоане, С., Лаза, А., Уррутксурту, М., и Ориве, Э.: Фитопланктон комплексы и их доминирующие пигменты в устье реки Нервион, Гидробиология, 549, 1–13, https://doi.org/10.1007/s10750-005-1736-6, 2005.
Ши, С. Л., Мари, Д., Жардилье, Л., Скэнлан, Д. Дж., и Воло, Д. : Группы без культурных представителей доминируют над эукариотами Пикофитопланктон олиготрофной юго-восточной части Тихого океана, Plos One, 4, e7657, https://doi.org/10.1371/journal.pone.0007657, 2009.
Синнинге Дамсте, Дж. С., Рампен, С., Ирен, В., Рупстра, К., Аббас, Б.,
Мюйзер, Г., и Схоутен, С.: Диатомовое происхождение длинноцепочечных диолов и
среднецепочечные гидроксиметилалканоаты, широко распространенные в четвертичных морских
отложения: Индикаторы питательной среды // Геохим.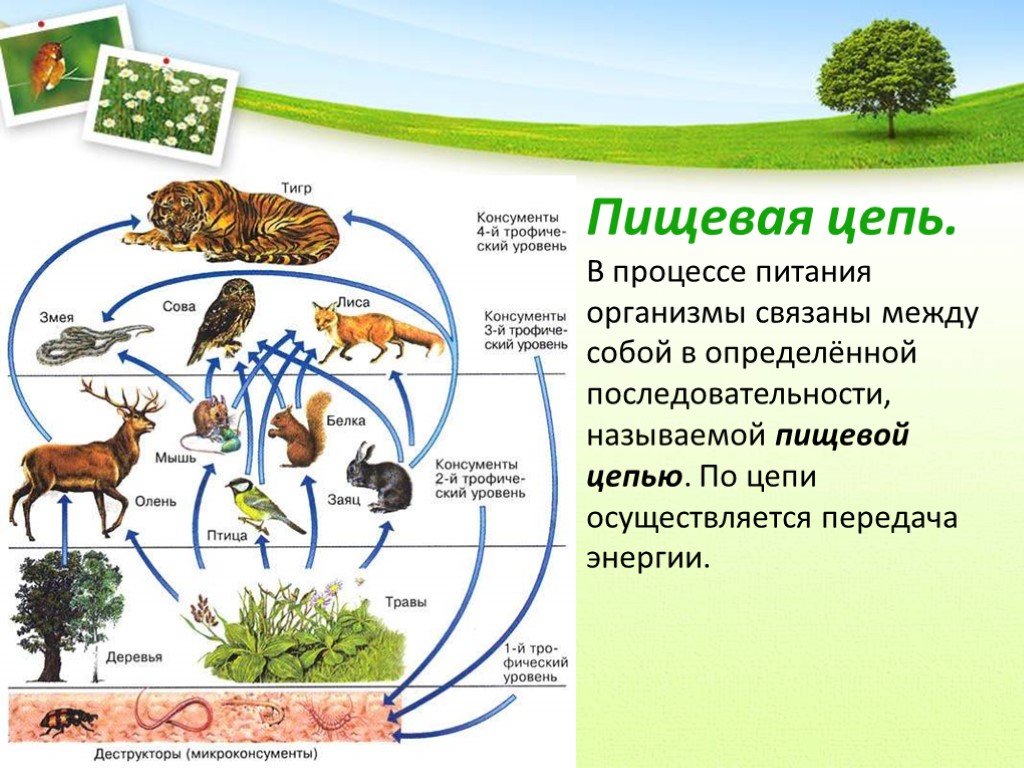 Космохим. Ак.,
67, 1339–1348, https://doi.org/10.1016/s0016-7037(02)01225-5, 2003.
Космохим. Ак.,
67, 1339–1348, https://doi.org/10.1016/s0016-7037(02)01225-5, 2003.
Слапета, Дж., Морейра, Д., и Лопес-Гарсия, П.: Распространенность протистов разнообразие: выводы из молекулярной экологии пресноводных эукариот, П. Рой. соц. Б-Биол. наук, 272, 2073–2081, https://doi.org/10.1098/rspb.2005.3195, 2005.
Speelman, E.N., Reichart, G.J., de Leeuw, J.W., Rijpstra, W.I.C., и Sinninghe Damste, JS: Биомаркеры липидов пресноводного папоротника Azolla и его ископаемый аналог из эоценового Северного Ледовитого океана, Org. Геохим., 40, оф. 628–637, https://doi.org/10.1016/j.orggeochem.2009.02.001, 2009.
Сток, А., Юргенс, К., Бунге, Дж., и Стоек, Т.: Разнообразие протистов в субкислородные и бескислородные воды Готландской впадины (Балтийское море) по данным Библиотеки клонов 18S рРНК, Aquat. микроб. экол., 55, 267–284, https://doi.org/10.3354/ame01301, 2009.
Стоек, Т., Бенке, А., Кристен, Р., Амарал-Зеттлер, Л., Родригес-Мора, М.
Дж., Чистосердов А.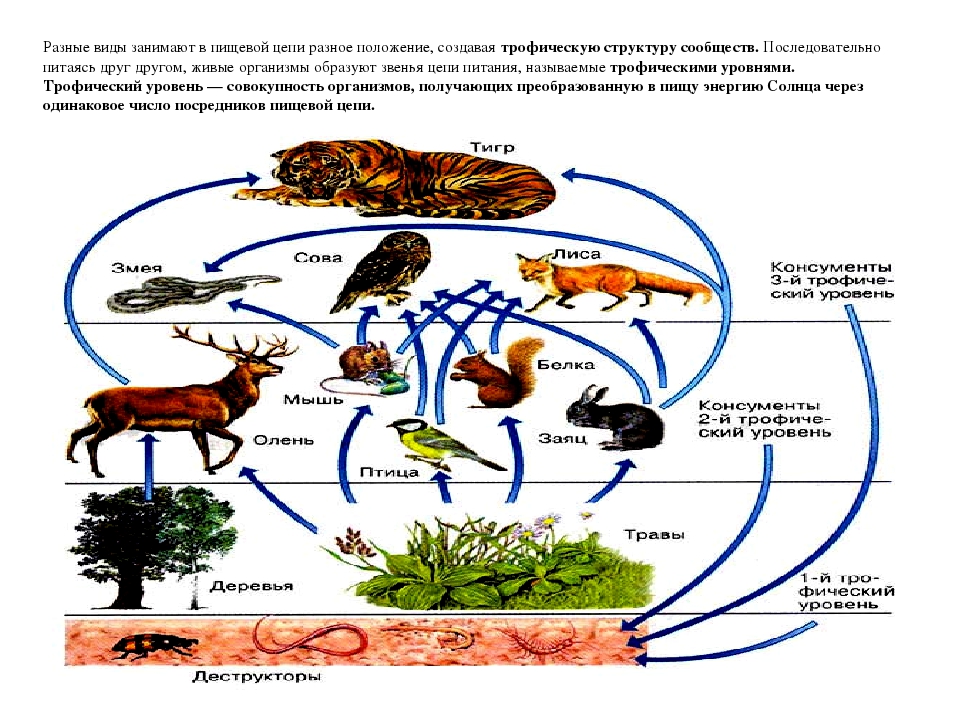 , Орси В., Эджкомб В. П. Массивно-параллельный тег
секвенирование показывает сложность анаэробных сообществ морских протистанов,
BMC Biol., 7, 72, https://doi.org/10.1186/1741-7007-7-72, 2009 г..
, Орси В., Эджкомб В. П. Массивно-параллельный тег
секвенирование показывает сложность анаэробных сообществ морских протистанов,
BMC Biol., 7, 72, https://doi.org/10.1186/1741-7007-7-72, 2009 г..
Стоек Т., Басс Д., Небель М., Кристен Р., Джонс М. Д. М., Брейнер Х. В. и Ричардс Т.А.: Параллельная метка множественной маркерной ДНК окружающей среды секвенирование выявляет очень сложное эукариотическое сообщество в морских бескислородных вода, мол. Экол., 19, 21–31, https://doi.org/10.1111/j.1365-294X.2009.04480.x, 2010.
Тегелаар, Э. В., Делеу, Дж. В., Деренн, С., и Ларжо, К.: Переоценка керогенообразования // Геохим. Космохим. Ак., 53, 3103–3106, https://doi.org/10.1016/0016-7037(89)
-9, 1989. Versteegh, GJM, Bosch, HJ, and De Leeuw, JW: Потенциал
палеоэкологическая информация о среднецепочечных диолах от С-24 до С-36, кетоолах
и гидроксижирные кислоты со средней длиной цепи; критический обзор, Org. Геохим., 27, оф.
1–13, https://doi.org/10.1016/s0146-6380(97)00063-6, 1997 г.
Versteegh, G.J.M., Jansen, J.H.F., De Leeuw, J.W., и Schneider, R. Р.: Диолы со средней длиной цепи и кетоолы в отложениях юго-восточной Атлантики: новый инструмент для прослеживание морских поверхностных водных масс?, Геохим. Космохим. Ак., 64, с. 1879 г.–1892, https://doi.org/10.1016/s0016-7037(99)00398-1, 2000.
Вильянуэва, Л., Бесселинг, М., Родриго-Гамис, М., Рампен, С. В., Вершурен, Д., и Синнинге Дамсте, Дж. С.: Потенциальные биологические источники цепные алкилдиолы в озерной системе, Org. Геохим., 68, 27–30, https://doi.org/10.1016/j.orggeochem.2014.01.001, 2014.
Фолькман, Дж. К., Барретт, С. М., Данстан, Г. А., и Джеффри, С. В.: C-30-C-32 Алкилдиолы и ненасыщенные спирты в микроводорослях класса Eustigmatophyceae, орг. геохим., 18, 131–138, https://doi.org/10.1016/0146-6380(92)
-v, 1992.
Wen, M. and Jetter, R.: Гидроксиальдегиды с очень длинной цепью из кутикулярного
воск хвои Taxus baccata, Фитохимия, 68, 2563–2569,
https://doi.org/10.